
www.jornaldaciencia.com.br
https://www.quora.com/profile/Sodr%C3%A9-Neto-1
1. https://www.academia.edu/39120976/Speciation_in_real_time_and_historical-archaeological_and_its_absence_in_geological_time
2. http://entropia.comunidades.net/teoria-do-neocatastrofismo-de-impactos-sodre-neto
3. http://ortomolecular.comunidades.net/a-cura-do-cancer-linfocitos-ativados-sodre-neto

TNI - TEORIA NEOCATASTROFISTA
DE IMPACTOS
Como Produzir Bilhões de anos em 30 Minutos e o Paradoxo da Estase Morfológica
Sodré Neto
HISTORIA DO ARTIGO
"Assustado com a reação busquei mais fontes que corroborassem com meu artigo para defendê-lo desta tempestade de críticas e um dos primeiros artigos que encontrei foi do grande cientista alemão Wolf-Ekkehard Lönnig e fiquei digamos aliviado ao ler quase as mesmas observações em "Dynamic Genomes" . Raciocinei que se tão grande cientista pensou as mesmas coisas eu não estaria tão absurdamente errado como a academia brasileira alardeava".

"Em 1944 (Simpson, 1944) fez uma meta-análise e concluiu a partir de 58 estudos publicados sobre padrões de especiação no registro fóssil que 71% apresentavam estase (e permanência) morfológica. A este respeito Michael Benton declarou: "parece claro, então, que a estase é comum, e isso não foi previsto a partir de estudos genéticos modernos" (Benton, 2009; Michael Oards, 2017), que preveem imensa variação de formas a partir do comportamento plástico dos seres vivos observados sobretudo desde Charles Darwin.
Por quase uma década mesmo sem ter o conhecimento de muitas destas pesquisas sobre a anomalia da "estase morfológica" , defendia o que chamava de "Paleontologia em T" , destacando o contraste entre os achados fósseis se repetindo em permanência de formas, com a altíssima variabilidade de formas recentes na "atualidade", e me baseei nesta época mais na diferença taxonômica e depois também no contraste entre fósseis repetidos em suas mesmas formas com seres vivos plásticos em seu comportamento usual (Lauvoué, 2009)
Anos mais tarde um cientista experiente em publicações (Everton Fernandes) me contactou para que publicássemos juntos algum dos muitos assuntos que me envolvi. Sugeri este tema (assunto que me motiva) com o título: "A Especiação em Tempo Real, em Tempo Histórico-Arqueológico e sua Ausência em Tempo Geocronológico" sendo publicado em inglês "Speciation in real time and historical-archaeological and its absence in geological time" publicado pela Academia Publishing sob DOI: 10.15413/ajsr.2017.0413 em maio de 2017. PDF com revisão em inglês
Durante a construção do artigo houveram varias descobertas e uma delas o Everton descobriu que, o que eu defendia fazia vários anos, como "Paleontologia em T", era chamado na literatura de "estase morfológica" que destaca mais um padrão fóssil de imobilidade de variação de formas, justificando-o, do que foca em contrastar as formas permanentes no registro fóssil com o comportamento não-permanente em mesmas formas na biodiversidade atual (super diversificada e se variando em formas (morfologicamente), até mesmo em mesmas especies quando sob quaisquer mudanças ambientais, pois "nenhum organismo ignora seu ambiente"(Ezard et e al, 2016).
De posse das palavras chaves "estase morfológica" foi bem mais fácil encontrar inúmeros outros artigos científicos tratando da questão até encontrar a expressão (contendo em si mesma) a flagrante e escandalosa admissão do "maior problema da evolução" (Mayer): PMS (Paradox of Morphological Stasis) "o paradoxo da estase morfológica" e são estas as palavras chaves do nosso antigo artigo.
No Brasil agora em 2018 teremos um candidato a presidência da república que também não se envergonha de apoiar tortura, ditadura militar, dar socos em público quase gratuitamente e expressar ódios aos pobres e gays, ele também fala que quer destruir o congresso, que democracia não funciona, defendendo a ditadura e já disse que vai começar uma revolução matando umas 30 mil pessoas!!! Mesmo assim ele é amado por muitos que o apelidaram carinhosamente de "mito"; analogamente assim vejo os artigos científicos antes da meta-análise de Simpson em 1944 e até hoje com as mais diversas revisões bibliográficas seguidas de mais justificativas para esta vergonhosa e escandalosa admissão do que admitem ser o paradoxo da estase morfológica.
A idéia do nosso artigo anterior era apenas demonstrar a anomalia das mudanças de formas (esperadas mas não achadas) e do padrão da repetição de formas e repetição taxonômica fóssil (mesmas espécies se repetindo até em estratos diferentes) como evidencia de sepultamento de seres básicos de todo planeta, surgidos prontos na coluna geológica.
Logicamente que isso tinha sérias implicâncias não somente desfavorecendo o suposto historico evolucionista das especies, como apoiava a idéia criacionista com 200 anos de altíssima rivalidade , de uma catástrofe mundial capaz de sepultar milhares de populações e por isso estarem repetidas em suas mesmas formas nos achados fosseis.
Quando a academia brasileira soube da publicação houve drástica reação . Ameaçaram a terceira autora de perder a bolsa de pós doutorado em evolução (era a segunda perseguição frontal que a pobre garota recebia) e mesmo sem respeitar o estado "grávido" dela, muito provavelmente a pressionaram para que solicitasse a remoção de seu nome, o que a revista se recusou fazê-lo. Na época tive então que declarar a isenção ao máximo de sua participação (até porque comigo não sei quantificar , mas acredito que foram poucas mesmo, não sei com o segundo autor) depois fizeram uma tempestade de protestos contra o artigo nas redes sociais, onde tentei quase em vão , discutir o assunto do artigo e chamar para o debate quem se dispusesse.
Podemos observar como destaco no artigo abaixo, que muitos cientistas tecem comentários a respeito deste ponto com extremo cuidado em suas publicações, agem como dando alfinetadas e não raras vezes usando frases , que se parecem irônicas, como julgo que foi o caso de Lauvoué ao declarar que "estudar o conservadorismo morfológico a longo prazo é difícil nos sistemas contemporâneos, porque poucas linhagens animais existentes são conhecidas por permanecerem morfologicamente estáticas em relação às escalas de tempo geológicas". (Lavoué et al, 2011).Isso se dá porque a academia mundial principalmente das áreas de geologia e biologia, é bem conhecida pelo seu comportamento repleto de ódio e repulsa para com artigos científicos criacionistas e do design inteligente (conforme um dos mais assistidos documentários do mundo "Expelled: No Intelligence Allowed"), o qual descreve dezenas de casos onde cientistas respeitados foram boicotados, mandados embora, prejudicados por apoiarem de alguma forma o criacionismo ou o design inteligente, e previsivelmente reagiu repudiando o artigo devido o caráter nitidamente a favor do "criacionismo".
Era até esperada esta reação por conta da nossa "afronta e audácia" de publicar dentro do terreno protegido e vigiado pelo que o Dr. Eberlin (um dos maiores bioquímicos da atualidade) chama de "patrulha darwinista", que deixou escapar a publicação em jornal internacional de idéias a favor do seu arqui-rival histórico e político (criacionismo) que desde o iluminismo tratou de considerar indistintamente separação política entre igreja e Estado (laicismo político) como válidos também na educação (laicismo educacional), tratando de simplesmente proibir assuntos como Deus em sala de aula e varrendo o que o pais da ciência chamavam de principio básico (Criador) para algo inexistente, criando assim, além de uma imensa prole de ateus , uma distância gigantesca entre o pensamento dos pais da ciência moderna e a própria ciência moderna, que agora, sob os moldes da camisa de força "naturalista" para assuntos religiosos, substituindo-os por mera ficção científica (sopa primordial, panspermia, biopoeise, etc) se tornando ciência moderna preconceituosa quase apenas em relação a temas relacionados a Deus.
Em menos de uma semana Houveram provavelmente dezenas de milhares de reações acadêmicas no Brasil e algumas internacionais como de uma autoridade britânica, quase todas sem conteúdo. Houve também uma reação do Mauricio Tuffani, ex chefe do caderno de ciência da Folha de São Paulo, que como observou Junior Eskelsen, demonstrou sequer compreender o título do artigo ao destacar que o mesmo se "apoiava" em "baraminology" assunto que foi apenas citado.
Na maioria das reações não houve apontamento ou demonstração de entendimento do assunto , houveram 3 ou 4 cientistas que se deram ao trabalho de debater numa página destinada a isso , mas a maioria teve como saída apenas atacá-lo por ser criacionista e por ter sido publicado numa revista da lista das possíveis "predatórias" apesar que o corpo editorial da mesma possuir boas notas B3, B4 conforme lista de 2017 pela agência oficial CAPES que apoia pesquisadores brasileiros. (outro detalhe que enquanto o site cita comportamentos de uma lista de revistas cientificas como sendo "possíveis" predatórias, os brasileiros já afirmaram logo)
Assustado com a reação busquei mais fontes que corroborassem com meu artigo para defendê-lo desta tempestade de críticas e um dos primeiros artigos que encontrei foi do grande cientista alemão (não poderia ser um brasileiro? ) Wolf-Ekkehard Lönnig e fiquei digamos aliviado ao ler quase as mesmas observações em "Dynamic Genomes" . Raciocinei que se tão grande cientista pensou as mesmas coisas eu não estaria tão absurdamente errado como a academia brasileira alardeava.
Encorajado a pesquisar mais busquei antecipar objeções, e dar resposta mais completa, abrangente e eficiente para satisfazer questões levantadas pelo mesmo artigo. Comecei fazendo acréscimos e revisões para antecipar as críticas, mas pouco tempo a revisão se transformou não somente num novo artigo, mas em uma teoria, com centenas de elementos novos ou mais desenvolvidos, centenas de artigos citados a mais, onde o ponto principal que era " o paradoxo da estase morfológica"observada como anomalia sem qualquer harmonia com a teoria de Darwin (observado por Darwin e antes dele), se tornou ponto lateral e coadjuvante junto com dezenas de outros, formando um novo modelo geocronológico de impactos que denominamos "Teoria Neocatastrofista de Impactos" com a qual percebemos que os diversos problemas atuais da historia da terra inclusive a PMS (Paradox of Morphological Stasis), funcionam neste novo modelo como peças de uma quebra-cabeças permitindo uma visão cada vez mais harmônica com a realidade.Recentemente encontrei experiente cientista seguindo mesma linha (Michael Oards), além dos muitos citados neste trabalho como o Dr. Walter Brown, Loening do Max Plank, Dr. Nahor Souza Junior que foi quem me deu os fundamentos da geologia catastrofista e o primeiro a falar para mim que os impactos provavelmente eram "o segredo" e foi a partir de um mega debate de 9000 réplicas e tréplicas, com 4 professores de geologia durante 1 ano e meio sob assistência do mesmo, que percebi um pouco que os impactos eram o ponto chave para a compreensão da historia da terra e sobretudo do fanerozoico, os quais complementam dentro de suas especialidades e pontos observados, a confirmação de que estamos apenas um pouco mais acima dos efeitos dos mesmos.
Por fim busquei e ainda busco uma centenas de revisores, críticos, colaboradores e co-autores entre os quais cito Hector Lutero, que foi quem percebeu geniosamente a hipótese de tendência de relação de diâmetro do bólido com idade (Figura 1), Dan de Barna, que me forneceu um capítulo inteiro de seu livro citandio o famoso cientista "Alvares" , Roberto Carlos Machado que forneceu relatos arqueológicos de catástrofes e tem pesquisado sobre cinturão de asteroides, e fui agregando alguns pontos importantes e revisando outros, podendo oferecer agora com uma certa expectativa positiva, uma previsão de trabalho ao público em geral, que mesmo possuindo diversos pontos que ainda necessitam de maiores estudos e demonstrações, ofereço para que a discussão do mesmo e sua exposição, possa melhorar ou criar novos projetos de pesquisa atrelados, que dignifiquem cada vez mais a ciência dos fatos, nossas descobertas , estes novos autores e suas novas ou antigas publicações, com novas observações e olhares ; e porque não mencionar também, para que dignifiquem sobretudo e sobre todos ao nosso Criador.
A todos uma boa leitura!
Sodré Neto
geologiaatual@gmail.com
31 975352798
https://sodregoncalves.no.comunidades.net/mudancas-morfologicas-em-tempo-geologico
TNI - TEORIA
NEOCATASTROFISTA DE
IMPACTOS
Como Produzir Bilhões de anos em 30 Minutos e o Paradoxo da Estase Morfológica
Sodré Neto
VIDEO EXPLICANDO O ARTIGO E COMO ACHAR DIAMANTES

TNI - CONEXÕES IMEDIATAS SEPARADAS POR ABSOLUTISMO
TNI - TEORIA NEOCATASTROFISTA DE IMPACTOS, CONEXÕES IMEDIATAS SEPARADAS POR GEOCRONOLOGIA ABSOLUTISTA![]()
Sodré Neto
geologiaatual@gmail.com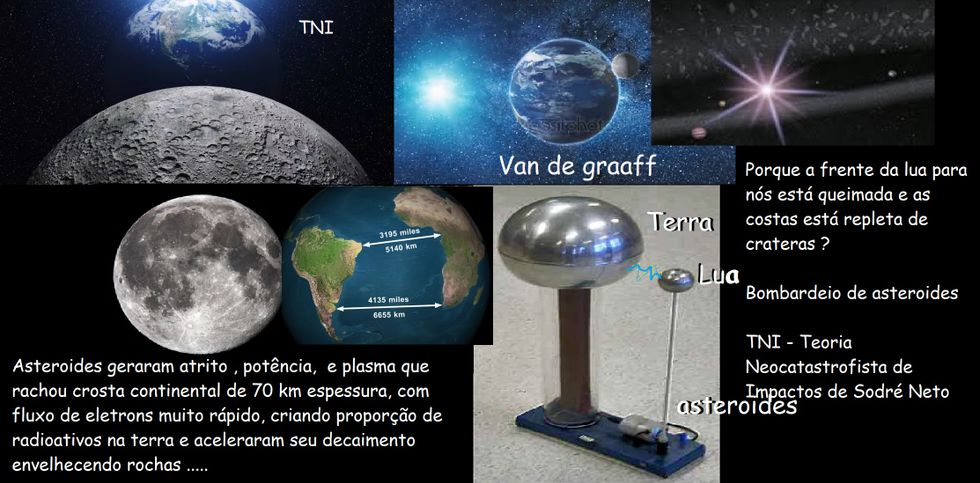
Basta clicar no link [discussão "x"] ao lado da afirmação, que você vai cair num tópico destinado a colaborações, comentários, críticas, sugestões e/ou refutações , no grupo TNI e outros de astronomia , geologia, paleontologia, biologia , etc..do facebook . Experimente!!!
|
EVIDÊNCIAS DUPLAS QUE UNEM ACONTECIMENTOS SEPARADOS PELO TEMPO |
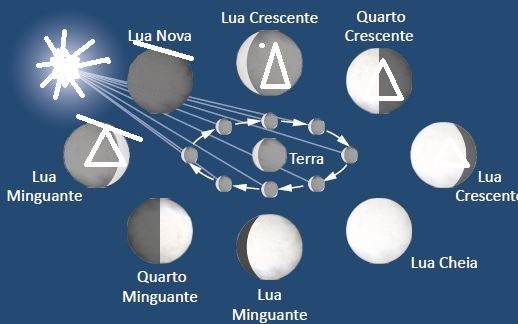
RESUMO - Conexões imediatas separadas pelo tempo se refere a fatos conectados um depois do outro imediatamente , mas devido as datações classificadas como absolutas , fazem com que as leituras paleontológicas, geológicas, astrofísicas e biológicas , estejam sendo interpretados como acontecimentos distantes no tempo um do outro. Preparamos um modelo neste trabalho que pretende resolver isso e para tanto iniciamos perguntas chaves que serão respondidas no decorrer do presente trabalho. São elas : 1) Porque a lua está queimada para nosso lado e repleta de 30.000 crateras do lado oposto? 2) Se podemos ver 30.000 crateras no maciço da lua, onde foi parar e porque sumiram 99,6 a 99,8% (50 a 100.000 crateras ) da terra? 3) Como se comportou o manto em face de uma chuva de asteroides? 4) Quando a crosta continental que sustenta a américa do sul e de outros continentes, de 70 km médios de espessura, estava unida a crosta continental da áfrica e outros continentes , era menor nosso planeta? 5) O manto engoliu asteroides e há evidências disso? 6) O sistema terra-lua com a terra tendo 4000 elementos químicos e lua maciça com apenas em torno de 100 elementos químicos, quando sujeitos a um bombardeio de asteroides , criaram uma tensão diferencial de carga e funcionaram como uma bola de Van de Graaff ? 7) A diferença de cargas poderia ter queimado o nosso lado da lua gerando fluxo de elétrons tão imenso que criou elementos radioativos e acelerou o decaimento "envelhecendo" rochas para terem aparência de "bilhões de anos" em poucos segundos ? 8) O bombardeio de asteroides estavam positivados pelo atrito e na terra aquecida muitos mergulhariam no cristalino aquecido pelos impactos anteriores; que efeito isso traria ? 9) Parte dos asteroides do bombardeio mergulharam no manto, no próprio magma exposto que estaria formando a crosta marinha ? 10) Na lua maciça apenas chocaram, e esta diferença entre lua e terra geraria gigantesco diferencial elétrico capaz de gerar imensa tensão entre os dois corpos? 11) Neste caso fabricaria um gigantesco plasma e curto circuito que cortaria a crosta da terra? 12) O que mais racharia uma crosta com espessura de 70 km senão um plasma criado com a altíssima amperagem criada por um bombardeio de asteroides no sistema terra-lua? 13) Este bombardeio foi recente ou faz muitos anos? 14) Deveria haver trilhões de evidências de um acidente recente na terra? Existem ? 15) Se existem e são claros o que "proibiu" a geologia de perceber o óbvio?
Toda a terra está repleta de sinais de gigantescas catástrofes ocorridas recentemente, estarei listando em torno de umas 50 perspectivas isócronas que atestam tal fato. Cito algumas como introdução:
1)Carbono 14 em quantidade datável , presente em rochas do fanerozoico, consideradas como tendo 300-500 milhões de anos, e também em diamantes incontamináveis incrustados nestas rochas, foram testados no laboratório de Los Álamos pelo geofísico Dr. John Baumgardner e equipe , publicaram em 2004, e revelaram que tais rochas são recentes e não podem possuir a idade de centena de milhões de anos e nem mesmo de mais de 70 mil anos. Ele tambem tem publicado novos modelos para comportamento do manto terrestre http://adsabs.harvard.edu/abs/2017AGUFMMR24B..01C 2) Trilhões de Pedras pontiagudas na terra revelam existir recentemente pois suas pontas estariam desgastadas caso fossem velhas. Num mesmo terreno encontramos uma ao lado de outra , uma arredondada e outra pontiaguda . Ora, a erosão que arredondou as arestas de uma de mesmo material no mesmo terreno não foi capaz de arredondar a outra ?Sua repetição nos estratos geológicos une sua idade recente umas as outras, além de revelar um desastre gigantesco recente que as fabricou. 3) Rochas pouco desgastadas por impactos de águas enérgicas em cachoeiras de vários terrenos considerados velhos, une as mesmas a um tempo recente e comum. 4) Repetição de 71% das formas fósseis sob a luz da observação evolutiva ou da forte influência que o ambiente exerce mudando as formas (morfologia) dos seres vivos, nos declara que esta repetição morfológica em "estase", permanente, de mesmas formas, apenas confirma que viveram sob um mesmo período e sob um mesmo ambiente, onde nossa observação do comportamento plástico dos seres vivos, condena a ideia que pertenceram a tempos distintos. A repetição de formas fósseis em 71% dos seres vivos (Simpson, 1944, Benton 2009) demonstra ainda o sepultamento de quase todas as populações de espécies na terra (pois se houvesse mudanças ambientais + tempo, nunca teríamos permanência de mesmas formas fósseis) 5) A meia-vida curta do DNA explicitado nas publicações do geneticista John C Sanford , junto com o geofísico John Baumgardner e outros, ao mesmo tempo que encurta a possibilidade de tempo dos seres vivos na terra, reúne todos os seres vivos a uma época recente. 6) A queda de grandes bólidos e seus efeitos elétricos criando plasmas tem o poder de destruir a confiança na "constância de decaimento" em sistema "fechado" e nos faz prever rochas "envelhecidas radiometricamente" pela aceleração dos ponteiros do relógio radiométrico como demonstra inúmeras tecnicas patenteadas de descontaminação usando aceleração de decaimento em sistemas de aceleração de partículas 7) A junção de acontecimentos separados pelo tempo , como a queda do Chicchulub tendo causado o Dekkan (Richards, 2015) nos impede de aceitar que tais acontecimentos unidos um ao outro, estejam separados por milhões de anos. 8) Tecidos moles de pequenos "bifes" endurecidos de tiranossauro-rex preservados nos impede de concluir que sua extinção foi a muito tempo, mas combina entre dezenas de evidências (9-73) que ela foi recente e não a 68 milhões de anos como a geocronologia convencional afirma. Tais observações fortalecem a proposta da hipótese de uma chuva de asteroides e meteoros a pouco tempo atrás e nos faz recomendar um esboço falseável para uma suposta confirmação ao pesquisar a) ângulo de alguns impactos b) Familia dos impactos (por exemplo, o chichulub tem autores que defendem que ele é da familia bapstina) c) Em havendo hipótese de ângulo comum com impactos na lua e na terra , podemos prever em que órbita os que não bateram a 4-5 mil anos atrás , estarão hoje? A resposta a esta pergunta a pesquisa responderá.
http://entropia.comunidades.net/evidencias-duplas-unindo-fatos-separados-no-tempo
Talvez deste quando deparamos com flagrantes achados de tecidos moles (minúsculos bifes com elasticidade endurecidos encontrados juntos com ossos com sua maior parte sem sofrerem permineralização e fossilização) de tiranossauro -rex, datados em "absurdos" chamados de "absolutos" 68 milhões de anos, refutados aqui e dezenas de outros como Triceratops horridus onde se diz (Armitage, 2013)
What is also not clear is how such biofilm structures could themselves survive the ravages of time, as once produced other microorganisms could begin to digest even these
O que também não está claro é como essas estruturas de biofilme poderiam sobreviver à deterioração do tempo, uma vez que outros microrganismos poderiam começar a digerir inclusive esses.
https://doi.org/10.1016/j.acthis.2013.01.001
Ou quando testamos mesmas rochas ao lado da outra com idades bem diferentes , ou estudamos correntes elétricas na queda de grandes asteroides , formando plasma e seus efeitos de aceleração de decaimento nuclear gerando consequente "envelhecimento" de rochas, que pode ocorrer na mente do pesquisador , que toda geocronologia está absurdamente e absolutamente errada e que os cientistas só não declaram isso com medo de enfrentar retaliação dos sacerdotes da doutrina ideológica que como religião substituta , domina com caneta de aço e perseguições aos cientistas "hereges", desde Darwin, a academia ainda hoje.
Veja no minuto 9:17-9:40 aparece o tecido sendo esticado e voltando!
Justificaremos abaixo inúmeros outros fatos, dezenas de perspectivas e centenas de razões, incluindo leis e hipóteses testadas e outras por testar, para propor a TNI (Teoria neocatastrofista de impactos) como ensaio isento da cronologia oficial dos fatos . Estas justificativas não somente sustentam este ensaio como exigem que desvencilhemos do absolutismo datacional que se tornou uma convenção na academia que exalta alguns pontos favoráveis e tenta colocar pra debaixo do tapete elefantes que discordam.
Nossa simulação inicia com a hipótese de um bombardeio lunar e terrestre. Na lua existem 30.000 crateras de impacto (astroblemas) de asteroides e meteoros, e na terra, temos apenas 200 astroblemas catalogados e 2000 em sites russos. Como a lua está muito próxima , e a terra possui campo de atracão mais forte, e diâmetro 3,67 maior que a lua, calcula-se que ela tenha recebido mais de 50 a 100.000 impactos que: 1) estão ainda por ser descobertos? 2) Foram engolidos devido o calor da crosta sofrendo efeito de plasmas elétricos gerados quando castigada por uma chuva de asteroides? 3) Haveria rastros de acreções?
Causas cósmicas para eventos terrestres parecem estar invadindo a geologia moderna. Neste artigo, 34 autores liderados pelo Dr. Edward J. Steele, apresenta um bombardeio de asteroides como causa da "explosão" cambriana; bem como considera bombardeamento de bólidos como estando presentes nos principais pontos de mudança geológico-evolucionaria da terra.. " major extinction-diversification evolutionary boundaries coincide .. cometary-bolide bombardment events" "Os principais limites evolutivos da extinção-diversificação coincidem com os eventos de bombardeamento por bólidos-cometas" ejsteele@cyo.edu.au https://www.sciencedirect.com/science/article/pii/S0079610718300798?via%3Dihub
Porém observamos que um bombardeio de asteroides não fabricaria "apenas" algumas camadas estratigráficas; e sim um conjunto de eventos tectônicos, envolvendo pluma de manto, elétricos, plasma, geoquimicos e não apenas sedimentares .
Entre muitas pesquisas e publicações catastrofistas, que parecem superar as uniformistas na geologia atual , outra publicação aponta para medição de 0.05% do tempo estimado anteriormente, para a formação das camadas do cambriano, combinando com a idéia catastrófica dos asteroides acima.
Berthault, G., Lalomov, A. V. and Tugarova, M. A. : “Reconstruction of paleolithodynamic formation conditions of Cambrian-Ordovician sandstones in the Northwestern Russian platform” – “Lithology and Mineral Resources, 2011, Volume 46, Number 1, 60-70” (Springer Publishing site)
Apesar do trabalho de Steele buscar justificativas para a panspermia,como fenômeno mais provável para inicio complexo da vida no lugar da velha sopa "morna" (Darwin, Miller, Oparin) primordial, [Discussão "complexidade da vida"], ele se reporta a ação cósmica, como alternativa natural mais provável, citando outros trabalhos como :
"explosão Cambriana e a escala de extinção em massa sugere a passagem do nosso Sistema Solar através de uma Nuvem Molecular Gigante desalojando múltiplos cometas da Nuvem de Oort para o interior do Sistema Solar, causando impactos na Terra ( Hoyle e Wickramasinghe, 1981). , 1993 )." ciclo) com impactos cometários coincidentes com eventos de extinção em massa ( Clube et al., 1996 ) e diversificações evolutivas ( Napier et al., 2007 ).
https://www.scopus.com/inward/record.url?eid=2-s2.0-37549068536&partnerID=10&rel=R3.0.0
https://www.scopus.com/inward/record.url?eid=2-s2.0-0039611522&partnerID=10&rel=R3.0.0
Uma de nossas previsões de 9 anos atrás foi recentemente publicada (Richards, Mark; 2015) referentes ao efeito mantélico do deccan na índia, causado pelo Chicxulub, onde comentamos a possibilidade de aquecimento da crosta durante o bombardeio de asteroides gerando efeitos diversos sobretudo elétricos, plasma, e como isso auxiliaria haver penetração do bólido, sua expressão no lado oposto (assim como vemos as costas e a frente da lua contendo um mar de basalto), acresções destes corpos na terra , aumentando sua matéria, inchando, inflacionando a terra (aumentando o dia de 18 horas pra 24 horas) e criando as placas tectônicas e a crosta marinha recente, lembrando que tal efeito se repetiria em todos os pontos além do dekkan no quadro abaixo: Tarbuck Lutgens - Earth An Introdution ti physical Geology. 11 edition. Pag. 449
Tarbuck Lutgens - Earth An Introdution ti physical Geology. 11 edition. Pag. 449
O Plasma de Torus 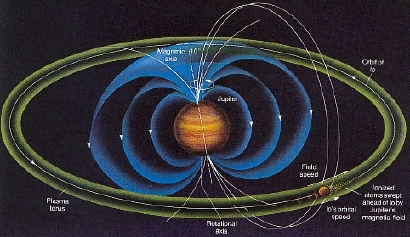
Em Jupiter parece que ocorre algo similar ao que poderia ter ocorrido na terra . Uma enorme corrente elétrica flui . "Aproximadamente 2 trilhões de watts de energia são gerados. A corrente segue as linhas do campo magnético até a superfície de Júpiter, onde cria raios na alta atmosfera. A primeira imagem em preto e branco do Telescópio Espacial Hubble mostra o tubo de fluxo" http://planetaryexploration-net.planetpatriot.net/jupiter/io/io_plasma_torus.html
Um fato curioso nos salta aos olhos quando avaliamos as 200 crateras da terra catalogadas . As "rochas do fanerozóico contêm 155 impactos e as rochas pre-cambrianas contêm apenas 27, ou 15% do total reportado" (Oard, Michael J., 2009). Justamente nas bacias sedimentares que "apagam" mais , temos uma proporção maior de crateras que nas camadas pre-cambrianas que "apagam" menos. Fora o fato que as rochas e terrenos pré-cambrianos, representam o testemunho de 4,5 bilhão de anos, ou 9 vezes mais o suposto tempo do fanerozoico (544 Ma).
Esperaríamos que terrenos de bacias sedimentares, devido sua dinâmica de receber sedimentos e fornecer sedimentos, guardassem na memoria geológica, menor numero de marcas de crateras, e perguntamos como pode as bacias guardarem mais memória, mesmo estando bem menos expostas e possuindo maior capacidade e dinâmica de apagamento das crateras?
Ainda dois pontos podemos observar, que por curto período de tempo, menos que 1/9 cada estrato, estiveram as camadas do fanerozoico expostas, enquanto as pre-cambrianas por bilhões de anos. por outro lado, raciocinamos, sob a perspectiva de chuva de asteroides, se as crateras só apareceriam nas partes mais "frias" sedimentares do planeta, uma vez que as rochas pre-cambrianas estariam aquecidas o suficiente para engolir os impactos.
Neste ponto buscamos uma solução . Estas crateras e seus efeitos estão separados pela geocronologia convencional , contudo, estudos ligados a bombardeamento, cinturões de asteroides, e envelhecimento de rochas pelos aspectos elétricos (plasma) , pressão, e aceleradores de partículas envolvidos num impacto, os quais envelheceriam rochas, nos fazem prever que uma historia geocronológica bem diferente da atual existe e que teremos que recorrer a diversas perspectivas datacionais cruzadas para compor uma historia mais próxima a real idade dos eventos. 
"Esta foto divulgada pelo Observatório Europeu do Sul (ESO) na terça-feira mostra a impressão de um artista do asteróide exilado 2004 EW95, o primeiro asteróide rico em carbono, confirmado no cinturão de Kuiper e uma relíquia do sistema solar primordial. Este curioso objeto provavelmente se formou no cinturão de asteroides entre Marte e Júpiter e deve ter sido transportado bilhões de quilômetros desde sua origem até sua atual residência no Cinturão de Kuiper, de acordo com o European Southern Observatory."https://arxiv.org/abs/1801.10163
Interpretaremos alguns dados, à luz de uma nova perspectiva que estamos propondo, que surgiu a partir do estudo de dados, primeiramente paleontológicos e evolutivos, que nos recomendaram (Sodré et al, 2017) pesquisar possibilidade de sepultamento de todas as populações de seres vivos ancestrais da terra devido a repetição de formas fósseis (PMS- paradox stasis morphologix) em 71% das espécies (Simpson, 1944; Benton, 2009), o que reclamaria a ação de um cinturão de asteroides.
A hipótese de chuva de asteroides tem sido discutida pelos astrofisicos:
"Foi hipotetizado que os impactores que criaram a maioria das crateras observáveis nas terras altas lunares antigas foram derivados do cinturão principal de asteróides de tal forma que preservaram sua distribuição de freqüência de tamanho". (Minton, 2014)
https://www.sciencedirect.com/science/article/pii/S0019103514005570
https://www.researchgate.net/publication/265015081_Re-examining_the_main_asteroid_belt_as_the_primary_source_of_ancient_lunar_craters
"It has been hypothesized that the impactors that created the majority of the observable craters on the ancient lunar highlands were derived from the main asteroid belt in such a way that preserved their size-frequency distribution".
"Foi hipotetizado que as antigas crateras das terras altas lunares foram formadas por impactos de uma população que era idêntica à do cinturão principal de asteróides , baseada nas similaridades entre a distribuição de tamanho-freqüência dos asteróides e a distribuição de freqüência-tamanho dos impactadores das terras altas lunares. usando uma relação de escala de tamanho da cratera " " Os impactadores lunares ... "têm uma distribuição de tamanho muito semelhante à dos asteróides próximos da Terra" ( Strom et al., 2005 ) https://www.scopus.com/record/display.uri?eid=2-s2.0-24944537842&origin=inward&txGid=861f289ad3679c29bb237560a9a851e3
http://dx.doi.org/10.1006/icar.2001.6608
https://www.sciencedirect.com/science/article/pii/S0019103501966084?via%3Dihub
Pesquisamos o que ocorreu enquanto parte do cinturão de asteroides, ejetado pelo movimento de Júpiter , não entrasse em órbita em Kuiper. Ele estaria "procurando" a sua estabilidade e órbita, e é neste contexto cosmológico que iniciamos nossa discussão. [discussão Asteroides de Marte a Kuiper] [discussão na comunidade "astronomia em debate"] . Tal procura implica em considerar a velocidade descomunal do cinturão ejetado , e o gigantesco caminho até encontrar sua órbita. Ao considerar este momento de instabilidade na trajetória, percebemos sua relação bem estreita com os bombardeios de asteroides que ocorreram na terra e na lua. Tsiganis et al., 2005 , Morbidelli et al., 2005 , Gomes et al., 2005 )
https://www.scopus.com/inward/record.url?eid=2-s2.0-19544378163&partnerID=10&rel=R3.0.0
https://www.sciencedirect.com/science/article/pii/S0019103509003029
https://doi.org/10.1038/nature03676
https://www.sciencedirect.com/science/article/pii/S0019103517303408
https://www.sciencedirect.com/science/article/pii/S0019103501966084?via%3Dihub
DATAÇÃO SEPARA EVENTOS E SUA AUSÊNCIA OS UNE
Assumindo que diversos bombardeios marcaram as extinções em massa, perguntamos se estes bombardeamentos estiveram ou não separados por grandes períodos de tempo, e quê evidencias sedimentares teríamos deste distanciamento, se a forma plano-paralela em que os estratos estão, apresentam inúmeras razões para que haja uma interpretação de formação conjunta, estratificando e separando elementos comuns desagregados, além de não apresentam quase nenhum vestígio de formações necessitadas de tempo para justificar intercalação de períodos entre uma camada e outra.
Perguntamos sobretudo qual segurança podemos ter nos métodos de datação radiométricos, chamados de "absolutos", (que é um termo anti-científico que lembra justamente o que o iluminismo combateu: O absolutismo dos reis empossados pela autoridade divina dos papas "infalíveis") .
Que segurança nos fornecem para termos pelo menos "a quase certeza" de que durante bilhões de anos, o decaimento radioativo esteve trabalhando constantemente? [Discussão plasma/sistemas e aceleração de decaimento na comunidade "loucos por Física"]
Condenamos esta premissa uniformista principalmente porque sabemos que certos ambientes elétricos como plasma, etc, a constância de decaimento, base do relógio radiométrico, não acontece, e pode ser acelerada produzindo rochas de bilhões de anos em apenas segundo(s).
Modelos propostos pelo Dr. Walter Brown citam " Dentro da crosta de granito no início do dilúvio, o efeito piezoelétrico gerou tensões suficientemente altas para iniciar os fluxos de plasma - colapsos elétricos - dentro da crosta e fusão que produziram novos elementos químicos - muitos radioativos."
http://www.creationscience.com/onlinebook/Radioactivity2.html#wp35970265
Na figura abaixo, percebemos uma pista nesta direção: Investigamos 200 crateras , seu diâmetro e a idade atribuída, e percebemos que os bólidos não apenas atingem a terra, mas também nossas "certezas" "absolutas" geocronológicas; Pois percebemos uma relação entre diâmetro dos bólidos e sua idade. Depois verificamos esta mesma relação na lua e outros planetas.
Figura 1 - Linha de aumento de idade relacionada ao diâmetro de bólidos . Sodré & Lutero, 2017

***Observamos que há uma linha de tendência em relação a dizer que numericamente diâmetros maiores dos bólidos possuem maior probabilidade de estarem relacionados a maiores idades, e numericamente bólidos pequenos ajuntados em torno de idades menores, possuem proporcionalmente probabilidade de estarem relacionados a idades menores .
DECAIMENTO RADIOATIVO, PRODUZINDO BILHÕES DE ANOS EM SEGUNDOS
Alta pressão e variação de decaimento
http://siba-ese.unisalento.it/index.php/psba5/article/view/18228
HAHN, H., BOBN, H. & KIM, J. (1976). Inquérito à Taxa de Perturbação da Queda Nuclear. Radiochimica Acta , 23 (1), pp. 23-37. Retirado 9 de abril de 2018, do doi: 10.1524 / ract.1976.23.1.23
Em 2009 a mesma idéia se repete
https://www.sciencedirect.com/science/article/pii/S0370269309001063
Decaimento alfa em ambientes de elétrons de densidade crescente: Do núcleo nu à matéria comprimida
https://link.springer.com/article/10.1140/epja/i2016-16032-3
https://www.sciencedirect.com/science/article/pii/S092765050900084X
http://www.creationscience.com/onlinebook/IntheBeginningTOC.html
Meia-vida de decaimento alfa de 221 Fr em diferentes ambientes
https://link.springer.com/article/10.1140/epja/i2007-10011-9
Problemas nucleares em q-plasma
http://www.scielo.br/scielo.php?pid=S0103-97332009000400006&script=sci_arttext
![]()

William A. Barker descreve seu pedido de patente de aceleração de decaimento radioativo (como método de descontaminação de materiais radioativos) ,em 31 dez. 1991, da seguinte maneira :
"De um modo geral, a comunidade científica acredita que a taxa de decaimento de um núcleo radioativo é imutável. No entanto, é possível alterar a taxa de decaimento alterando o ambiente do emissor.... Desta forma, a taxa de decaimento da radioactividade dos materiais é grandemente acelerada e os materiais são assim descontaminados a uma velocidade muito mais rápida do que o normal. O estímulo pode ser aplicado aos materiais radioativos, colocando esses materiais dentro da esfera ou terminal de um gerador de Van de Graaff onde eles são submetidos ao potencial elétrico do gerador, como na faixa de 50 kilovolts a 500 kilovolts, para em pelo menos um período de 30 minutos ou mais. A presente invenção baseia-se no facto de a taxa de decomposição de materiais radioactivos poderem ser aceleradas ou reforçadas e assim ser controlada por um estímulo, tal como um potencial electrostático aplicado. Esse potencial, por exemplo, é incorporado na equação de tunelamento mecânico quântico para o coeficiente de transmissão T * T, incluindo uma energia potencial adicional"
Muitos outros trabalhos e pedidos de patente para métodos de aceleração de decaimento radioativo e descontaminação de materiais estão descritos na literatura (An Kinderewitscg, 2003; Gorodezki, 2005) . O aparato de Willian Parker citado precisou de 50-500 kilovolts para gerar aceleração de decaimento e descontaminar assim em apenas 30 minutos, mas nos falta equipamentos para medir os efeitos de milhões de kilovolts e o que geraria a queda de apenas 1 grande bólido. E também como simular qual seria as consequências, em termos de aceleração de decaimento e envelhecimento de rochas , diversos bólidos caindo, eletrificando, aquecendo e fragmentando a crosta terrestre, criando plumas mantélicas, etc.
As experiências com fusão nuclear em inúmeros testes e projetos como de equipamentos de tokamaks , de que , através de sistemas de plasma e temperatura, poder não somente aumentar decaimento mas até alterar núcleo de elementos estáveis (Bosch, 1999; Lee, 2008; Hesslow L et al, 2017; Izumi et al, 2016; Zhang et al, 2016; Xie et al, 2014) , e os efeitos de plasmas e outros aceleradores de partículas durante a queda de grandes bólidos (dos quais temos catalogado apenas em torno de 0,2%), tendo nós mesmos dado uma contribuição relevante para tendência de decaimento acelerado em relação ao diâmetro do bólido (Figura 1), tudo isso nos assegura que podemos simular aqui uma interpretação isenta da dependência tradicional da geocronologia devido esta não poder mais estar (pelo menos “absoluta”) diante de tais testes e fatos e de muitas outras perspectivas datacionais anacrônicas, sem necessitar portanto de tais inúmeras justificativas ad hoc, podendo simular uma interpretação com os dados como eles simplesmente são e estão.
A interpretação isenta da geocronologia convencional e tradicionalmente ensinada desde dois últimos séculos, economiza por assim dizer, uma série de malabarismos justificadores de anomalias anacrônicas que pululam nas descobertas científicas, mas ela trará novos desafios como por exemplo, se não temos este tempo todo distanciando a queda de bólido do outro, então poderíamos nos aproximar mais de estudos da NASA de múltiplos impactos na história da terra ? (Spray, 1998; Donald R. Lowe, 2004; Bunch, 2012; Witke JH, 2013; Kennett, 2015). A evasão de gases e o isolamento dos raios solares pelas grossas nuvens de fumaça esfriariam rapidamente a superfície da terra, criando a glaciação, em cima de uma crosta fervendo logo abaixo dos continentes e das águas em movimento ? Poderia a queda de grandes bólidos envelhecer rochas pela aceleração de decaimento radioativo? poderia transformar alguns elementos estáveis em instáveis arrancando nêutrons de seus núcleos?
A queda de maiores bólidos representam potencial de aceleração maior de partículas e de possível até mesmo surgimento de elementos instáveis (Brown, 2013). As poucas exceções a esta tendencia verificada , podem ter explicações em relação ao terreno do impacto se apresentaria ou não amortecimento do impacto, e o mesmo vale para bólidos de diâmetros pequenos se o terreno impactado teria gerado ou não maiores fatores de aceleração de partículas.
A queda de um bólido maior implica em maior temperatura conjugada a maior efeito plasma conjugado a maior efeito piezoelétrico e a maior onda sonora; todos estes efeitos são aceleradores de partículas e , em graus cada vez mais elevados, são mais capazes de arrancar neutrons até de elementos estáveis, bem como estes neutrôns soltos se agregar a outros elementos criando instabilidade por neutrons a mais e a menos nos novos elementos radioativos gerados.
Se possuem tais potenciais de aceleração de decaimento maior, logo, bólidos maiores teriam a tendencia de ter maiores idades como revela o gráfico e bólidos menores , menores idades como demonstra o gráfico.
Ao observar as idades de rochas defrontamos com as rochas mais velhas do planeta no cinturão de rochas verdes Nuvvuagittuq próxima ao arco de Nastapoka no litoral litoral da costa sudeste da Baía de Hudson , no Canadá . Isso combina perfeitamente com nossas observações das ações de impactos (Beals, 1968; Goodings, 1992; Bleeker, 2004) no envelhecimento de rochas. 
Por teoria entendemos o conjunto de hipóteses testadas, que passaram por testes de falseabilidade (Popper) associada a hipóteses ainda em estudo e pesquisa, que agregando um conjunto formado por diversas perspectivas de diversas áreas distintas (neste caso, astronômicas, físicas, químicas, genéticas, biológicas, paleontológicas, geológicas, arqueológicas e outras áreas), criam por assim dizer um modelo teórico que pode ou não ser mais comunicável com a realidade dos fatos.
Tanto em ciência, como em qualquer área da vida, é uma boa prática evitar dogmatismos impondo nossas idéias, e tentar separar ao máximo aquilo que são dados, daquilo que é nossa interpretação deles. As muitas religiões e opiniões distintas que se agregam em torno de interpretações de textos sagrados, tem seu espelho na ciência em torno da interpretação dos dados emitidos nas publicações científicas, e o mesmo cuidado e problemas que aqueles enfrentam, a ciência também enfrenta, até porque, em última análise, todos estamos em grande medida perdidos, tateando em busca de sentido para todas as coisas, sejam mais físicas e mais testáveis, sejam mais abstratas e mais conjecturais . Lembrando que também foi (Mayr, 2006) que sugere uma divisão entre evolução biológica mais funcional, de evolução histórica mais conjectural.
Efeitos elétricos como plasma tem sido objeto de estudo de muitos cientistas (ex: Peratt, Anthony L. https://www.scopus.com/authid/detail.uri?authorId=7003878552 ) .Chegamos ao conhecimento destes e das mesmas idéias , por meio de investigação dos dados, e não influenciados pelos mesmos. De forma que os dados nos uniram .
Há estudos que mostram similaridades entre os cinturões http://iopscience.iop.org/article/10.3847/2041-8213/aab3dc/meta
https://pubs.geoscienceworld.org/sepm/gsabulletin/article/127/11-12/1507/126064
Pesquisa relacionada em https://scholar.google.com.br/scholar?hl=pt-BR&as_sdt=0%2C5&q=mercury+impact+asteroid+plume+large+province+igneous&btnG=
https://scholar.google.com/scholar?hl=es&as_sdt=0%2C5&q=+asteroid+impact+mercury&btnG=

“All quartz-rich rocks (quartzites, granites, gneisses, mylonites) did show [statistically significant] piezoelectric effects when stressed.” J. R. Bishop, “Piezoelectric Effects in Quartz-Rich Rocks,” Tectonophysics, Vol. 77, 20 August 1981, p. 297. Alta pressão e variação de decaimento HAHN, H., BOBN, H. & KIM, J. (1976). Inquérito à Taxa de Perturbação da Queda Nuclear. Radiochimica Acta , 23 (1), pp. 23-37. Retirado 9 de abril de 2018, do doi: 10.1524 / ract.1976.23.1.23 Em 2009 a mesma idéia se repete Decaimento alfa em ambientes de elétrons de densidade crescente: Do núcleo nu à matéria comprimida https://www.sciencedirect.com/…/artic…/pii/S092765050900084X Meia-vida de decaimento alfa de 221 Fr em diferentes ambientes Problemas nucleares em q-plasma Se relativizarmos portanto tais constantes radioativas e universais, podemos mudar ou pelo menos tornar incertas certas "verdades" cientificas; e isso será muito bom para pelo menos salvar a ciência do dogmatismo quanto a historia do universo e da terra; Mas acima disto , diante de tantas contradições anacrônicas. as vezes bem flagrantes como datar tecidos moles em 68 milhões de anos(para não enumerar mais de 1000 outros casos) , destacamos que isso nos permite testar outros modelos de historia com os fatos para verificar quais hipóteses e teorias são mais comunicáveis com os mesmos, o que iremos fazer neste trabalho. Existem duas teorias em disputa na mentalidade de todos nós. Que a terra foi planejada, organizada e arquitetada para a vida ou que tudo ocorreu por acidentes cósmicos que por sorte geraram as condições necessárias para que diversos códigos genéticos contendo informação super inteligente atuassem e criassem nosso bio-sistema. "Even the largest and oldest known terrestrial impact structures
Alguns poucos desses dezenas de milhares de impactos ainda não identificados são expostos em sites como na russia http://labmpg.sscc.ru/impact/ e menos ainda são considerados pela criteriosa comunidade cientifica http://www.passc. Já no livro texto de geologia da USP "Decifrando a Terra" , percebemos um reflexo do que ocorre no mundo onde geólogos estão abolindo uma visão uniformista da historia da terra devido os acontecimentos passados não terem pares comparados aos atuais, restando a permanência apenas do atualismo assumindo mesmas leis no passado, porém não mesmos acontecimentos. O Dr. Baker cita que "a doutrina do uniformitarismo demonstraram há muito tempo que eram excessivamente restritivas na prática científica. O último deve ser relegado apenas ao interesse histórico no progresso das idéias" (Baker, 1998) Baker, Victor R. (1 de janeiro de 1998). «Catastrophism and uniformitarianism: logical roots and current relevance in geology». Geological Society, London, Special Publications (em inglês). 143 (1): 171–182. ISSN 0305-8719. doi:10.1144/GSL.SP.1998.143.01.15. Disponível em http://sp.lyellcollection.org/content/143/1/171.short em 16/12/2017 A razão desta mudança é exatamente o contraste de magnitude dos acontecimentos geológicos do passado com os atuais, e por isso podemos contemplar a terra e verificar facilmente como certas "construções" geológicas necessitaram de imensa e inimaginável energia catastrófica (muitas de magnitude e abrangência global em seus efeitos) para poder realizá-la. Uma pista em direção a mudança brusca do planeta se encontra no contraste verificado entre o padrão morfológico gigantesco fóssil comparado aos seus respectivos descendentes de hoje (e mesmo daqueles animais e plantas gigantescas que foram extintos), nos revela que este planeta na época destes fósseis era diferente e muito mais adaptável a vida, favorecendo expressões fenópticas maiores e para algumas espécies (como répteis) consequentemente mais longevas . As raízes da camada denominada carbonífero impressionam; para poderem crescer mais precisam se alimentar mais e terras sem pedregulhos propiciam tal crescimento e alimentação mais abundante . Nós pegamos uma cenoura e plantamos algumas em terra peneirada e outras em terra com pedregulhos e podemos testar este fato. Ora, a terra não poderia ter tantos pedregulhos para produzir as raízes do carbonífero, e seu ambiente deveria ser bem diferente do atual para abrigar animais gigantes e ainda permitir que existissem. O que aconteceu na terra que a encheu de pedregulhos e mudou drasticamente seu ambiente a ponto de podermos contrastar o mundo fóssil do atual sem gradualismos que justifiquem tal mudança drástica e contrastante? Goodings, CR & Brookfield, ME, 1992.Movimentos transcorrentes proterozóicos ao longo do linfonodo Kapuskasing (Província Superior, Canadá) e sua relação com as estruturas circundantes. Earth-Science Reviews, 32: 147-185. «Geoscience meets the four horsemen?: Tracking the rise of neocatastrophism». Global and Planetary Change (em inglês). 74 (1): 43–48. 1 de outubro de 2010. ISSN 0921-8181. doi:10.1016/j.gloplacha.2010.07.005 O uniformitarismo, apesar de ser uma ferramenta importante e até essencial para compreendermos os eventos geológicos e alguns até tentarmos reproduzir, foi substituído e até certo ponto minimizado por atualismo geológico que admite que apenas mesmas leis operavam "uniformemente" no passado. Esta mudança se deve ao fato de se perceber que os processos geológicos do passado não se repetirem na sua magnitude no presente. (Marrine, 2010) lista 7 fatores que tem contribuído para transformar a perspectiva interpretativa dos mais atualizados trabalhos geológicos: Ele intitula como: "1) o aumento da geociência aplicada; 2) epistemologia geológica herdada; (3) interação disciplinar e difusão de idéias das ciências planetárias para a terra; (4) o advento das técnicas de namoramento radiométrico; (5) a revolução das comunicações; (6) webometria e a busca por geociências de alto impacto; e (7) estruturas culturais populares" . Marrine, Nick (1 de outubro de 2010). «Geoscience meets the four horsemen?: Tracking the rise of neocatastrophism». Global and Planetary Change (em inglês). 74 (1): 43–48. ISSN 0921-8181. doi:10.1016/j.gloplacha.2010.07.005 https://www.sciencedirect.com/science/article/pii/S092181811000158X Esta velha e agora renascida leitura interpretativa geológica percebe os fenômenos catastróficos do passado como causadores de grandes alterações globais na superfície terrestre suplantando interpretações antes quase exclusivamente uniformistas e gradualistas . Os fenômenos catastróficos, em especial os de alta energia e consequências globais, como impactos de grandes asteroides, estão cada vez mais na mente do observador em geologia. A inserção de impactos e seus efeitos na coluna geológica é um fato recente e na opinião de alguns geólogos deveria causar uma revolução muito maior que simplesmente deixar tudo como estava sendo considerado, como se os impactos fossem apenas um detalhe sazonal nos milhões de anos de formação sedimentar , mas percebemos que uma vez que impactos estão atrelados a formações sedimentares e estas se relacionam umas as outras , nos parece mais provável que esta inserção advogue um novo modelo de historia geológica da terra. A questão datacional parece conter uma série de reclamações anacrônicas que tem tornado cada vez mais insustentável manter a geocronologia como ponto "resolvido" . Contudo, defender a abolição de seu absolutismo nos coloca na posição de inclusive ter que namorar a hipótese de chuva de asteroides como vem ocorrendo em centenas de publicações sobre "multiplos impactos", "impactos binarios" , "bombardeamento de impactos" . (Ormö, 2014; Hasller, 2001; Glikson, 2004) Se fosse possível a abolição do absolutismo radiométrico abriríamos a possibilidade de que um bombardeio continuo de impactos tiveram estreita ligação com todas as formações sedimentares entre o cambriano/ediacara e camadas denominadas do pleistosceno. Outro ponto que raciocinamos em cima do artigo de (Ormö, 2014) é que se há fartas evidencias de impactos binários oriundos de uma mesma colisão, múltiplos impactos e asteroides caindo em cima de outro ainda não resfriado, sugerem cenário cada vez mais próximo de um bombardeio.
The surprisingly found anomaly that there are no significant morphological changes (including those towards phylogenies and disparate morphologies) in the fossil record to confirm and validate the evolutionary tree of ancestry totally common and / or at least represent the major environmental changes occurring over millions of years, is called in the literature "paradox of morphological stasis" (PMS) and in this work we will demonstrate its relation with the geochronology and stratigraphy under the impact of asteroids occurred in the earth. O ponto inicial e central no nosso artigo anterior (Sodré et all, 2017) era o paradoxo da estase morfológica que no entendimento de muitos é um forte testemunho do sepultamento de uma só geração de seres vivos na terra. Mas como foi possível existir este sepulatamento do planeta? Neste presente artigo visualizamos dezenas de aspectos que corroboram com a PMS (Paradox of Morphological Stasis) para formar um quadro harmônico com este novo modelo que denominamos TNI :Teoria Neocatastrofista de Impactos. Devido se perceber que estase morfológica no registro fóssil, seguido de repetição de mesmas espécies, representar o sepultamento de uma população, em uma geração, em um só momento, defendemos que qualquer interpretação que avance um milímetro sequer desta afirmação, contrariará tudo que conhecemos sobre evolução e plasticidade adaptativa das espécies ao longo do tempo, pois terá que defender o impossível: "estase morfológica" e permanência de formas, mesmo sob a pressão ambiental de diversas catástrofes ocorridas nos milhões de anos . O padrão chamado por "paradoxo da estase morfológica" no registro fóssil, (PMS - paradox os morphological stasis), representado por espécies com pouca modificação morfológica se repetindo em camadas sucessivas, quando comparamos se 1) representa espécies (em seu comportamento evolutivo) intercaladas por milhões de anos, que neste caso, teriam que estar muito modificadas morfologicamente, tanto mesmas espécies (que mudam morfologicamente em mudanças ambientais) e muito mais espécies que sofreram processo de especiação, ou se 2) representa um sepultamento de várias populações da terra se repetindo morfologicamente em estratos sedimentares (que neste caso teriam que estar sendo formados rapidamente e sucessivamente), é uma parte essencial do que tem estado em disputa durante os 2 últimos séculos de discussão sobre a origem das espécies, na dramática disputa entre criacionismo e darwinismo quanto ao melhor modelo de história de nossas origens. Devido raramente se observar que espécies descendentes não modificam , também raramente deveria se observar tal comportamento estável morfologicamente, nas respectivas ascendentes (ancestrais filogenéticos) da mesma morfologia. Isso nos tem inclinado a considerar com parcimônia a opção 2 como extremamente mais comunicável com a PMS. As interpretações que buscam uma interpretação do PMS sem confrontar o paradigma atual geocronológico e as chamadas datações absolutas, acabam se submetendo a um tipo de absolutismo que os obriga mais a justificarem a anomalia que a dialogarem com os dados. Neste trabalho indicamos uma saída "libertadora" deste absolutismo, apresentando evidências de aceleração de decaimento radioativo por ocasião da queda de bólidos e o consequente "envelhecimento" de rochas durante um muito provável bombardeamento de asteroides na terra. Temos que enxergar o fato de haver conflito de opinões e enfrentar o fato do pesquisador em geral não poder contrariar a tradição acadêmica ( establishment scientific) e seus respectivos chefes de departamento de pesquisa , e o resultado disso é que quase tudo que lemos em livros oficiais didáticos e publicações em revistas respeitáveis sobre evolução e plasticidade adaptativa das espécies ao longo do tempo, se comportam justificando a PMS classificando-o como "uma anomalia desafiadora" , sendo estes adjetivos talvêz seu limite e o máximo que os pesquisadores podem expressar para não queimarem seus nomes nas modernas fogueiras de "hereges" modernos. O darwinismo observável e estritamente biológico nos leva a esta conclusão sumária totalmente contra as previsões (não estáticas morfologicamente) dos seres vivos intercalados por supostos milhões de anos no passado (darwinismo histórico), ao mesmo tempo, perguntamos que opção haveria para explicar a PMS e um sepultamento simultâneo da população de toda a terra com diversos estratos geológicos? Em 1944 uma meta-análise concluiu a partir de 58 estudos publicados sobre padrões de especiação no registro fóssil que 71% apresentavam estase morfológica. A este respeito Michael Benton declarou: "parece claro, então, que a estase é comum, e isso não foi previsto a partir de estudos genéticos modernos" Simpson, GG (1944). Tempo e Modo na evolução. Columbia University Press. Nova Iorque, p. 203. Benton, Michael e David Harper (2009) Introdução à Paleobiologia e ao Fossil Record New York: John Wiley & Sons, pp. 123-124. PERSPECTIVAS QUE CONCORDAM COM A HIPÓTESE DE BOMBARDEAMENTO DE IMPACTOS Levantamos aqui a hipótese de uma chuva e bombardeamento de asteroides e percebemos que esta hipótese se comunica de forma econômica com diversos ítens harmônicos entre si , e através da mesma , poderemos justificar o PMS como um sepultamento de uma geração da população do mundo; sendo fortalecida por mais de dezenas de itens isócronos e interligados , que como peças de quebra-cabeça se harmonizam . Alguns citamos como necessitados de maiores pesquisas e outros comentamos e listamos como:
|









Colisão de Íons Pesados
Há diversas publicações de impactos binários , múltiplos e teve um achado recente na Austrália do impacto ter caído em cima do outro antes do mesmo esfriar ....O impacto cai gera altíssima eletrificação em seguida outro cai e no encontro dos efeitos ocorre colisão de íons pesados .
https://www.sprace.org.br/
https://www.sciencedirect.com/
https://de.wikipedia.org/wiki/
https://www.if.ufrgs.br/
E acabei entrando mais a respeito daquele experimento ALICE LHC que o Avelino me falou
" A maior parte das colisões de partículas realizada pelo LHC é entre dois prótons. Entretanto, também são feitas as colisões envolvendo íons pesados, em particular o chumbo. A diferença é que, ao usar um íon pesado, após a colisão, apenas por um instante, é formado o plasma de quark e glúons (QGP). Esse meio é o que os físicos acreditam ser similar ao do universo frações de segundos após o Big Bang."
http://griper.if.usp.br/node/
"A força de um buraco negro está diretamente relacionada com a quantidade de matéria (ou energia) que ele tem. Há físicos que dizem que a geração de buracos negros microscópicos dentro dos tubos do LHC é possível, embora nenhum tenha sido observado até agora. Entretanto, mesmo se eles fossem criados, sua força seria proporcional à energia das colisões – equivalente à energia de uma mosca batendo suas asas. Nenhum buraco negro criado no LHC, portanto, teria energia para engolir nada ao seu redor.
Além disso, sabemos que buracos negros perdem matéria através da emissão de energia por um processo descrito por Stephen Hawking. Qualquer buraco negro que não consegue engolir nada ao seu redor, como os que seriam produzidos pelo LHC, iriam desaparecer tão rápido que não poderiam nem mesmo ser detectados diretamente – sua presença seria notada devido ao seu decaimento, tipo de rastro que ele deixaria" https://www.sprace.org.br/
A poeira cósmica e o Iridium
Precisamos aqui separar asteroides de outros corpos de asteroides de corpos semelhantes ao terrestre. Naqueles veremos o que Alvarez observou em relação ao iridium :
http://science.sciencemag.org/
PDF http://science.sciencemag.
Nos outros poderemos interpretar como causadares da sedimentação de quase todo fanerozoico, onde mediana ordem de deposição poderia ser explicada pela forte dinâmica marinha.
O LADO OCULTO DA LUA REPLETO DE CRATERAS
Perguntamos se o lado que fica exposto para nós da lua não seria consequência da chuva de asteroides que recebeu em seu lado oculto..
27 dias, 7 horas e 43 minutos se repetem exatamente e sincronicamente (na rotação em torno de si) e (na revolução da lua em torno da terra), tendo seu lado sempre voltado para a terra e seu lado oposto para objetos que atacassem a terra, sempre exposto, que no caso estaria exposto especialmente num momento em que houve uma chuva de asteroides.
No vídeo abaixo se faz uma simulação de formação da lua
Em geral quando ocorre o confronto de uma única vez de dois corpos, o despedaçamento ocorre formando blocos maiores e centenas de milhares de blocos menores , tendo poucos jogados para bem mais longe que outro (resultado da tensão num ponto). Esta imagem se harmoniza com o presente estudo , pois observamos :
1. O lado escuro e mais "queimado" seriam manifestações mantélicas do que ocorreu no lado oposto?
2. Os efeitos plasma, piezoelétricos , ondas sonoras e aceleradores de partícula vão dar a impressão de que estas quedas foram mais antigas (Na lua onde não temos a dinâmica do mar criando estratos "moles" (não litificados) esta relação (observada por Hector Lutero Siman quando discorríamos o assunto) fica bem mais evidente; observe que a queda dos grandes bólidos lunares são considerados os mais "antigos" .
3. Esta imagem explicara casos mais esporádicos de meteoros mais próximos viajando ainda no espaço alguns provavelmente oriundos de uma mesma colisão.
O Triângulo da Lua como evidencia de bombardeamento de impactos 
Existe uma linha como que formando um triângulo na lua que sugere bombardeamento de um belt fino de asteroides. Na medida que ela se movimentava no percurso deles, ela ia sendo impactada pelo primeiro na ponta do triângulo que ia se alargando na medida que a lua se adentrava no belt de cinturão....em sua largura.
Isso pode nos dar o ângulo dos impactos que vieram para a terra e confirmarmos tanto a falta devido a intercepção na lua, quanto os que caíram.
Excelente pesquisa sugere este triânugulo em forma de fatia de pizza. Uma hipótese falseável e altamente forte para evidenciar de uma vez por todas os grande bombardeamento que a terra sofreu.

Diamantes Ausentes na Lua e Presente na Terra
Há registros de diamantes no olho de crateras na terra, mas isso é ausente na lua(XXXX)mesmo possuindo mesmas rochas formadas durante a queda de asteroides. Na terra se observa que a maioria dos diamantes vem do manto,(xxxx)mas como já vimos , tem havido uma relação do impacto em relação ao seu efeito do lado oposto; Na hipótese de uma chuva de asteroides e aquecimento da crosta a ponta de deixar que o bólido penetrasse a terra justificaríamos neste ponto o "desaparecimento" de 100.000 crateras previstas para a terra bem como a presença de diamantes aqui em contraste com sua ausência na lua sob semelhantes eventos e condições. Diamantes do manto seriam também formados por esta perspectiva da queda de bólidos numa terra contendo material carbônico para, se transformar em diamantes no manto, ou se formarem na hora do impacto e serem simplesmente transportados na hora do impacto para o manto.
Formação Rápida e Sucessiva das Camadas sedimentares entre o Cambriano e o Pleistoceno
Desenvolvi uma disputa sob ajuda do Dr. Nahor Souza Junior, durante 1 ano e meio, entre agosto de 2009 e dezembro de 2010, numa espécie de banca virtual, debatendo com 4 conceituados professores de geologia do Brasil, sobre se havia ou não aspectos sedimentares que demonstrassem algum tempo de grande intervalo entre a formação do cambriano e o pleistoceno. Depois de 9000 réplicas e tréplicas , não houve uma única evidência de intervalo grande de tempo que pudesse ser apresentada sem contestação. Já o contrário disso, tivemos inúmeras evidências sem nenhuma contestação até hoje as quais defendem esta formação sedimentar contendo aspectos catastróficos, rápidos, de abrangência e magnitude global, em diversas análises mostrando que em um momento, tempo curto, meses e anos, o planeta foi castigado por uma sucessão de varias catástrofes formadoras de camadas sedimentares todas acontecendo logo após a outra ou em conjunto.
Nesta época estávamos defendendo apenas um dos aspectos ou consequências destas catástrofes relatado em centenas de fontes da arqueologia comum e que a maioria dos pais da geologia moderna antes do darwinismo, defendiam ser o dilúvio bíblico. Contudo , entendemos hoje que o "tal" diluvio dos pais da geologia e de centenas de fontes arqueológicas , seria apenas uma das consequências de uma chuva de asteroides.
As quatrilhões de pedras despedaçadas expostas no planeta e outras quintilhões debaixo da terra, onde um bom percentual apresenta estarem roliças devido grandes movimentos , outras tantas ainda apresentam arestas sem estarem desgastadas (ou com pouco desgastes e ainda obtusas revelando pouco desgaste natural que existiriam caso houvesse muito tempo) . Tais pedras evidenciam um cenário de imensa catástrofe; ocorridas a pouco tempo pois que estão juntas umas as outras. Outras rochas estão exatamente debaixo de fortes impactos de águas sem que apresentem erosões de milhões de anos, mas de pouquíssimos milhares de anos, revelando pouco tempo pelo principio de sabedoria milenar chamado pelos índios de "Itararé" na língua indígena quer dizer "água mole em pedra dura tanto bate até que fura".
A Geological Society of America publicou um artigo
COFFIN, HAROLD G (1 de julho de 1971). «Vertical Flotation of Horsetails (Equisetum): Geological Implications». GSA Bulletin (em inglês). 82 (7). ISSN 0016-7606. doi:10.1130/0016-7606(1971)82[2019:VFOHEG]2.0.CO;2
«QUALIFICATIONS AND EXPERIENCE OF». origins.swau.edu. Consultado em 29 de novembro de 2017
comunidades.net. «Bacia do Rio Columbia - Implicações Cronológicas». www.sodregoncalves.rede.comunidades.net. Consultado em 29 de novembro de 2017
O Dr. Scott W. Hassler e o Dr. Bruce M. Simonson (Hassler, 2001) , ao defenderem a existência de múltiplos impactos formadores de camadas sedimentares próximas, propõe "isso como um modelo para que os trabalhadores possam testar em outras camadas de esférulas para ver se o modelo possui aplicabilidade geral. Isso pode ser mais difícil nos depósitos de Fanerozoico; a prevalência da bioturbação, uma vez que o Neoproterozóico pode obscurecer as estruturas mecânicas dos depósitos de impacto".

Outros trabalhos que seguem mesma linha sugerem que micro-esférulas são padrões para rastrear formações sedimentares associadas com grandes ondas e tsunamis, no trabalho de (Bunch, 2012) vemos a admissão de que um terço do planeta possui tais evidencias.

Bunch, Ted E.; Hermes, Robert E.; Moore, Andrew M.T.; Kennett, Douglas J.; Weaver, James C.; Wittke, James H.; DeCarli, Paul S.; Bischoff, James L.; Hillman, Gordon C. (10 de julho de 2012). «Very high-temperature impact melt products as evidence for cosmic airbursts and impacts 12,900 years ago». Proceedings of the National Academy of Sciences of the United States of America. 109 (28): E1903–E1912. ISSN 0027-8424. PMID 22711809. doi:10.1073/pnas.1204453109
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3396500/Neste contexto de micro-esférulas observa o Dr. Prasad, que "observamos uma diminuição da intensidade e do número de colisões dentro da ejetada com o aumento da distância" (Prasad et al., 2010 Meteoritics & Planetary Science 45, 990-1006 (2010).
Padrão de Surgimento de Formas Prontas no Registro Fóssil
Se a geocronologia de idades de bilhões de anos não pode ser afirmada com toda a pretensa certeza absoluta , então teríamos sérias consequencias estratigráficas e de impactos , pois encurtaríamos o tempo das sedimentações, e as colocaríamos como sucessivas e rápidas como temos fartas evidências disso como o proprio fato de estarem em plano paralelo sem erosões inter-camadas que representassem desgaste no tempo, e ajuntaríamos cada vez mais os impactos para mais perto um do outro, aproximando da idéia de publicações que sugerem chuva de asteroides e múltiplos impactos na terra como uma possibilidade de resposta para explicar o que fendeu violentamente a crosta continental de média de 70 kilometros de espessura, e formou a maioria dos cenários extremamente catastróficos que observamos na terra , bem como uma lista infinda de acontecimentos que requereriam grandes magnitudes globais de acontecimentos, como chegar a produzir altíssimas e inimagináveis taxas sedimentares (Sadler, 1981) que alcançasse os continentes e sepultasse vivo animais gigantes continentais completos. O máximo que encontramos hoje, de animais de grande porte, são pedaços mais resistentes sendo fossilizados.
Na lua existem 30.000 marcas de crateras (astroblemas) de grandes e pequenos asteroides e meteoros, e na terra, temos apenas 200 astroblemas. Como a lua está muito próxima , e a terra possui campo de atracão mais forte, se deduz que, sendo seu diâmetro , 3,67 maior que a lua, ela tenha recebido no mínimo 4 x 30.000 asteroides, ou seja, mais de 100.000 impactos foram erodidos, encobertos, e/ou estão ainda por ser descobertos na terra. Ou no dizer de Zellner:
"Novos dados orbitais de alta resolução de missões lunares recentes, resolução melhorada e sensibilidade da instrumentação analítica, desenvolvimento de novas técnicas analíticas para aquisição de idades para amostras lunares, reavaliação de dados da literatura e modelos dinâmicos atualizados de evolução do sistema solar que levam em consideração essas novas observações levaram a novas interpretações do início do bombardeamento da Lua (e por procuração, a Terra)" (Zellner, 1 de setembro de 2017).
Os trabalhos que sugerem aspectos de proximidade temporal entre a queda de um bólido e outro estão cada vez mais onipresentes na literatura, (Schmitz, 2001; Ormö, 2014, Philipp R Heck, 2017; ) , na revista Nature encontramos (Ormö, 2014) onde vemos por exemplo até mesmo proximidade sob a ótica da datação aceita:

«Figure 2: Paleogeography of Baltica and neighboring cratons at the time of the increased cosmic bombardment following the ~470 Ma asteroid breakup event, and timeline for the related meteorite falls (black dot and line) as well as known craters (red dots).». www.nature.com (em inglês). Consultado em 8 de janeiro de 2018
"Approximately 470 million years ago one of the largest cosmic catastrophes occurred in our solar system since the accretion of the planets. A 200-km large asteroid was disrupted by a collision in the Main Asteroid Belt, which spawned fragments into Earth crossing orbits. This had tremendous consequences for the meteorite production and cratering rate during several millions of years following the event. The 7.5-km wide Lockne crater, central Sweden, is known to be a member of this family. We here provide evidence that Lockne and its nearby companion, the 0.7-km diameter, contemporaneous, Målingen crater, formed by the impact of a binary, presumably ‘rubble pile’ asteroid. This newly discovered crater doublet provides a unique reference for impacts by combined, and poorly consolidated projectiles, as well as for the development of binary asteroids". (Ormö, 2014)
"A maioria dos meteoritos que caem hoje são os condritos comuns tipo H e L, mas os principais asteróides do cinto melhor posicionados para entregar meteoritos são os condritos LL 1,2. Isso sugere que o fluxo atual de meteoritos é dominado por fragmentos de eventos recentes de destruição de asteróides 3,4 e, portanto, não é representativo em prazos mais longos (100-Myr). Aqui apresentamos a primeira reconstrução da composição do fluxo de meteoritos de fundo na Terra em tais escalas de tempo. Do calcário que se formou cerca de um milhão de anos antes da separação do corpo parental L-chondrite 466 Myr atrás, recuperamos minerais relict a partir de micrometeoritos grosseiros. Por análises elementares e isotópicas de oxigênio, mostramos que antes de 466 Myr atrás, os achondritos de diferentes fontes asteroidais tinham abundâncias semelhantes ou maiores do que os condritos comuns. Os achondritos primitivos, tais como lodranitos e acapulcoites, juntamente com achondritos não agrupados relacionados, constituíram ~ 15-34% do fluxo em comparação com apenas ~ 0,45% hoje "
Meteoritos raros comuns no período ordovícico (PDF Download Available). Available from: https://www.researchgate.net/publication/312647580_Rare_meteorites_common_in_the_Ordovician_period [accessed Nov 21 2017].
Most meteorites that fall today are H and L type ordinary chondrites, yet the main belt asteroids best positioned to deliver meteorites are LL chondrites 1,2 . This suggests that the current meteorite flux is dominated by fragments from recent asteroid breakup events 3,4 and therefore is not representative over longer (100-Myr) timescales. Here we present the first reconstruction of the composition of the background meteorite flux to Earth on such timescales. From limestone that formed about one million years before the breakup of the L-chondrite parent body 466 Myr ago, we have recovered relict minerals from coarse micrometeorites. By elemental and oxygen-isotopic analyses, we show that before 466 Myr ago, achondrites from different asteroidal sources had similar or higher abundances than ordinary chondrites. The primitive achondrites, such as lodranites and acapulcoites, together with related ungrouped achondrites, made up ~15–34% of the flux compared with only ~0.45% today
Rare meteorites common in the Ordovician period (PDF Download Available). Available from: https://www.researchgate.net/publication/312647580_Rare_meteorites_common_in_the_Ordovician_period [accessed Nov 21 2017].
Ormö, Jens; Sturkell, Erik; Alwmark, Carl; Melosh, Jay (23 de outubro de 2014). «First known Terrestrial Impact of a Binary Asteroid from a Main Belt Breakup Event». Scientific Reports (em inglês). 4 (1). ISSN 2045-2322. doi:10.1038/srep06724 «Figure 2: Paleogeography of Baltica and neighboring cratons at the time of the increased cosmic bombardment following the ~470 Ma asteroid breakup event, and timeline for the related meteorite falls (black dot and line) as well as known craters (red dots).». www.nature.com (em inglês). Consultado em 8 de janeiro de 2018 Glikson, A.Y.; Allen, C.; Vickers, J. «Multiple 3.47-Ga-old asteroid impact fallout units, Pilbara Craton, Western Australia☆». Earth and Planetary Science Letters. 221 (1-4): 383–396. doi:10.1016/s0012-821x(04)00104-9
Como a Espessa Crosta poderia Rachar?
A crosta continental possui média de 70 km de espessura. Para que ela se fendesse, de forma violenta como foi (como podemos ver na plataforma da américa do sul e na plataforma da áfrica , onde existem debaixo da costa marítma, gigantescas erupções semelhantes aquelas da bacia da Guanabara no Rio de Janeiro- Brazil, que deveria estar também como as milhares de outras debaixo de água, mas por algum motivo ainda misterioso e não explicado, está soerguida. O mistério do porquê que apenas este pedaço da plataforma estaria soerguido com uma grande depressão ao centro , rodeada de grandes erupções vulcânicas de altíssimas escalas sugere uma imensa catástrofe como fonte causadora de tal formação ígnea. Nestas imagens abaixo unimos estudos feitos na rússia com semelhanças a bacia da guanabara-RJ. Estas semelhanças requerem maiores estudos , mas tentam vincular a separação violenta entre áfrica e américa, a diversos quadros geomorfológicos que provavelmente podem ter relação com impactos .
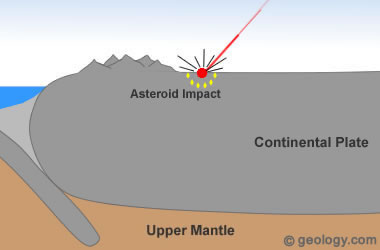
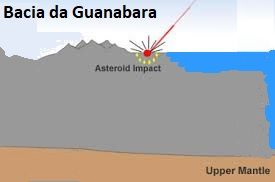
Bacia da Guanabara- RJ


�Popigai Crater no norte da Sibéria, na Rússia. http://geology.com/articles/diamonds-from-coal/ .Diagram from Guidebook 27, "Geology of Big Bend Ranch State Park, Texas" by C. D. Henry. Bacia da Guanabara - RJ - Brasil (Imagem de Popigal adaptada). Imagem CBERS da Baía de Guanabara (Fonte: INPE) - http://geologiamarinha.blogspot.com.br/2010/03/
Paradoxo da Estase Morfológica abrindo 8 janelas
Na pesquisa científica, a parcimônia é a escolha econômica de justificativas para uma observação, buscando assim a explicação mais simples e otimizada possível e na maioria das vezes, ela é considerada a melhor maneira de julgar uma hipótese (Courtney, 2008). Apresentamos neste trabalho, o que julgamos ser o caminho mais econômico e próximo de enxergar o paradoxo da estase morfológica (PMS) no registro fóssil, “que além de ser algo inesperado é ainda mais dramático porque:
"estudar o conservadorismo morfológico a longo prazo é difícil nos sistemas contemporâneos, porque poucas linhagens animais existentes são conhecidas por permanecerem morfologicamente estáticas em relação às escalas de tempo geológicas". (Lavoué et al, 2011).
O mesmo fato é observado por Peter Williamson, professor de Geologia da Universidade de Harvard, ao sugerir que o neodarwinismo tem falhado em não conseguir explicar as descontinuidades sistemáticas no registro fóssil:
"o principal problema é a estase morfológica. Uma teoria é somente tão boa quanto o são suas predições, e o neodarwinismo convencional, que alega ser uma explicação abrangente do processo evolutivo, falhou por não predizer a ampla estase morfológica, agora reconhecida como um dos aspectos mais impressionantes do registro fóssil." (Williamson, 1981, p.214).
Por muito tempo, na biologia evolutiva, o PMS tem confrontado a “evolução-fato” (mais experimental e observável) com a evolução histórica (mais conjectural e deduzida), onde a parte histórica, deduzida , mais conjectural e dependente da geocronologia de alta escala de tempo, apresenta um dos maiores problemas da teoria sintética da evolução (Mayr, 2002; Futuyama, 2010; Voge, 2016; Lonning, 2004), e isso tem levado pesquisadores a desenvolverem inúmeras justificativas, às vezes de extrema complexidade, às vezes extremamente distantes dos fatos materiais, dos dados concretos da paleontologia baseada em dados (Sepkoski, 2013), para tentarem harmonizar a falta de evolução e a falta de mudanças morfológicas, estacionadas em estase no registro fóssil , surpreendentemente presente durante supostos imensos períodos geocronológicos, o que realmente, pelo que conhecemos de evolução ocorrendo até em tempo real e a plasticidade responsiva dos seres vivos a mudanças no decorrer de pouco tempo, seria um completo absurdo, muito mais que uma simples anomalia, uma verdadeira aberração epistemológica.
A maioria dos trabalhos que defrontam com o PMS, também acumulam novas soluções, e outros apresentam soluções em estudo de casos declarados como quase “exclusivos” ou “extraordinários” (Lavoué, 2011), dando a entender que para o resto dos seres vivos, ou para casos não extraordinários, fica sem solução. A partir de observações de grande possibilidade de erro na geocronologia pela aceleração de decaimento provocado por plasmas , altas temperaturas geoquímicas (Kennett, 2015), inclusive locais de impacto que são ricos em diamantes devido o choque ter produzido altíssima temperatura e pressão para produzir os mesmos, e outros efeitos aceleradores de partículas (piezoelétricos e de ondas sonoras), por ocasião da queda de grandes bólidos (Figura1), referendando assim a possibilidade de uma interpretação dos dados, de forma direta e sem necessidade de justificativas, devido isenção da geocronologia tradicional que nos impõe uma necessidade de justificar a evolução ocorrendo mesmo sem dados morfológicos para tal, e também isenta da antiga idéia de Cuvier (1769-1832) que permanece praticamente constante até hoje na moderna teoria da evolução sintética , para explicar parciais sucessões de faunas estratigráficas, ignorando que já existe modernas propostas substitutivas de modelos de separação estratigráfica na literatura (Minoletti, 2009; Dilly et al, 2015: Berthaut, 1986, 1988, 2002, 2004, 2010, 2011, 2012, 2013, 2014; Lalomov, 2007, 2013; Julien, P.Y, 1993) que evocam, depois de análise dos dados, que uma revisão geocronológica e estratigráfica é necessária e se demonstra ser uma alternativa muito pertinente para se lidar com praticamente todos os problemas levantados pelo PMS, de forma parcimoniosa, simples, econômica e mais comunicável com fatos evolutivos , sedimentares, consequencias sedimentares e estratigráficas das catástrofes de impactos, e com outras perspectivas datacionais anacrônicas sempre necessitando de calibrações, ajustes e hipóteses justificadoras.
Por muito tempo a biologia evolutiva tem proposto diversas soluções para PMS e padrões fósseis ligados a ela. Em uma leitura rápida dos dados materiais poderíamos perceber que:
1) Mecanismos evolutivos estão ausentes no registro fóssil, pelo menos materialmente, devido a pobreza taxonômica de 250 a 300 mil espécies num mar de estimativas de haver trilhões de amostras fósseis no planeta
2) Igualdade técnica para números de padrões corporais de fósseis, quando comparados à imensa biodiversidade atual de estimativas de 8 a 100 milhões de espécies, revelam não somente a evolução totalmente parada em relação a diversificação no registro fóssil, como contrastadamente em rítmo acelerado hoje, mas mesmo assim acelerada, ocorrendo com limites numéricos de quase mesmo número de padrões corporais fósseis;
3) Estase morfológica revela também ambientes constantes e/ou segregados, onde os seres até mesmo de mesmo genótipo, teriam que viver sem mudanças e quaisquer pressões ambientais, nutricionais, e inclusive, sem tempo para que previsíveis mudanças morfológicas ocorressem no decorrer de tão longo tempo, para conseguir manter tal padrão morfológico;
4) Repetição de mesmas espécies fósseis em estratos distintos, como evidencía de sepultamento de grande taxa sedimentar capaz de formar diversos extratos , de uma mesma população aparecendo em estratos distintos , e abandonamos a idéia de formas intercaladas por milhões de anos pelos dados que demonstram que praticamente todas as espécies mudam morfologicamente com o tempo (exceto casos raríssimos);
5) O surgimento de formas de vida prontas como padrão no registro fóssil representa o que chamamos de catástrofe inédita segregadora e estratificadora de espécies , pois se este padrão pronto se repete , isso sugere, pela propria perspectiva e percepção evolucionária, algo não ocorrido antes. O padrão de surgimento pronto nos fala de algo que sepultou e fossilizou os seres prontos e se refuta a idéia de várias catástrofes de Cuvier atualizadas nas várias extinções em massa da teoria da evolução sintética moderna, dando lugar a uma catástrofe estraficadora contendo repetição de mesmas espécies em estratos diferentes (SRABURC - Standard of Ready Ancestors Buried in Unprecedented Catastrophism).
6) Surgimento pronto também possui ligação com o polêmico assunto da “complexidade irredutível” que já desafiava por meio de milhares de estudos bioquímicos da célula e sistemas de vida, sobre a impossibilidade de existência e sobrevivência de seres e sistemas sem que estivessem prontos (Behe, MJ, 1997, 2009; Khun, 2012, Looning, 2005) "O genoma do Octopus mostra um nível impressionante de complexidade com 33.000 genes codificadores de proteínas mais do que está presente no Homo sapiens" ( Albertin et al., 2015 ) https://www.scopus.com/record/display.uri?eid=2-s2.0-84939497927&origin=inward&txGid=fd980a234dd9d88ee593849b82cfa0ed .
Uma revisão crítica de Por que o neodarwinismo não funciona , por Fred Hoyle & Chandra Wickramasinghe
https://academic.oup.com/biolinnean/article/26/4/401/2670750 observa aspectos em favor da panspermia e contra a síntese evolutiva-genética como insustentável.
(e independente daquilo que isso evoca a ciência não se proíbe de evocar nada, senão que deve se ater aos fatos) . Estas observações nos fala também que:
7) um cenário rápido de sepultamento estratificador de grande parte da biodiversidade, algo condizente com rápidas, catastróficas e altas taxas sedimentares (Sadler, 1981) verificadas nas espessuras, larguras, extensões e pacotes únicos de camadas sedimentares e ígneas, onde o aumento de diversidade de sepultados (fósseis) seguido de queda coordenada de diminuição de proporção de fósseis, poderia espelhar também movimentos estratigráficos nos transportes nas transgressões e regressões marinhas e não necessariamente histórias de aumento de diversidade seguidas de extinções em massa. Esta interpretação dos dados estaria totalmente dependente de que a geocronologia de alta escala de tempo nas datações sobretudo das camadas sedimentares , chamadas de “absolutas”, estivessem totalmente erradas e que a antiga idéia de Cuvier (1769-1832) de sucessão e revolução faunísticas, intercaladas por muito tempo, e suas atualizações na moderna teoria sintética da evolução, na verdade, seria apenas um momento na história geológica onde diversas estratificações estiveram sendo construidas contendo repetição de espécies de uma mesma população. E por fim;
8) justificamos a recomendação de novos estudos ligados ás possíveis falhas da geocronologia convencional por meio de estudo de patentes de métodos de aceleração de decaimento radioativo, envelhecendo rochas em bilhões de anos em apenas 30 minutos, e observações de comportamento de plasmas atômicos aceleradores de decaimento radioativo, muito provavelmente gerados na terra por ocasião da queda de grandes bólidos, como capazes não somente de acelerar decaimento radioativo, como de fusão nuclear e/ou modificação nuclear em elementos estáveis.
O Surgimento sem antecedentes Evolucionários
O surgimento de formas de vida prontas, como padrão no registro fóssil, se comunica com idéia de impactos que gerariam grandes erosões e sedimentação atuando em conjunto com grandes transgressões e regressões marinhas, deixando rastros de camadas de sedimentos sobretudo nas bacias da terra. Os padrões de surgimento pronto, podem ser interpretados como uma consequência de gigantescas catastrofes associadas que ocorreram , em relação a vida na terra , de forma inédita, que teria que formar vários estratos contendo amostras de diversos seres vivos prontos nunca "surgidos" evolutivamente em outros estratos. Estudos nesta direção de múltiplos impactos vislumbram tais deposições e estratificações como :
"A deposição de S1, S2 e S3 foi amplamente influenciada por ondas e / ou correntes interpretadas para representar tsunamis gerados por impacto, e S1 e S2 mostram várias camadas graduadas que indicam a passagem de dois ou mais trens de ondas. Esses tsunamis podem ter promovido a mistura dentro de um oceano globalmente estratificado.." (Donald, 2004)
Este padrão descarta a presença de mecanismos evolutivos anteriores para as primeiras formas de vida, pois se um ou outro surgissem, alternando com antecedentes evolutivos e outros sem antecedentes, poderíamos até relevar este ponto e considerá-lo acidental devido uma série de justificativas (erosões, raridade fóssil, saltos evolutivos, etc.), mas estes gerariam fatores estocásticos e não regularidades padrões nos estratos, e o fato deste acontecimento ser padrão nos impõe imensa carga de ter que admitir que formas prontas simplesmente aconteceram na terra e não possuem sustentação, pelo menos material, dados, nos fósseis, para dizerem que foram evoluídas, ou que erosões e raridade fóssil estocásticas formaria este padrão em diversos estratos, etc. A opção de entender que estas formas prontas representariam uma imensa catástrofe sepultando pela primeira vez toda a vida na terra, se comunica com o que podemos chamar de "padrão de ancestrais prontos sepultados em diversos estratos em inédita e recente catastrofe" (SRABVSURC - standard of ready ancestors buried in various strata in unprecedented and recent catastrophe Catastrophism ). Sem contar com o fato da disputa que milhares de cientistas mais ligados a bioquímica das células fazem de que sistemas de seres vivos em geral precisam estar prontos devido necessitarem de “complexidade irredutíveis” (Behe, MJ, 1997, 2009; Khun, 2012, Looning, 2005) para poderem existir , se adaptar, sobreviver e evoluir .
O Dr. Kjetil L. Voge nos enviou sua publicação recentemente na qual se faz uma ampla revisão bibliográfica do problema da PMS (Voge, 2016). Inúmeros trabalhos correlatos passam a tocar o problema apresentando alguma solução para a anomalia surpreendente da estase morfológica ocorrendo em calculados Ma (milhões de anos). Todos parecem não duvidarem, nem questionarem com respeito aos problemas da "intocável" geocronologia e suas datações “absolutas”, nem da estratigrafia convencional, mas buscam dentro destes modelos, criar justificativas das mais diversas para a PMS.
Mas este não é um caso raro onde a geocronologia e estratigrafia impõe justificativas para dados anacrônicos, observamos que muitos outros pontos da paleontologia passam pela mesma situação, desde a sabedoria milenar dos indios ao pronunciarem a palavra "itararé" (água mole em pedra dura tanto bate até que fura" ) onde vemos trilhões de rochas recebendo impacto sem sofrerem erosões compatíveis a muito tempo, até pontos que vão estar ainda mais frontalmente destacando o anacronismo , como a datação de tecidos moles enrijecidos com borras amarronzadas de hemoglobina datados em extraordinários 60 a 120 milhões de anos. Um completo absurdo científico que prefere justificar e especular sobre super-poderes misteriosos da proteína e tecidos moles de preservarem por tanto tempo, que questionar o absolutismo datacional, apesar que estudos recentes estão desbancando tais possibilidades de preservação (Saitta, 2017) . Mas em geral os absurdos são tantos que para que este artigo não se transforme num livro, escolhemos apenas três exemplos bastante flagrantes , na Tabela 2 abaixo, citando uma amostra de ínfima quantidade proporcional de artigos relacionados, todos buscando encontrar saídas do tipo “ad hoc” para anomalias surpreendentes e anacrônicas. Destaco que alguns destes artigos parecem até ironizar o problema de forma a poderem ser publicados, sem confrontar diretamente o paradigma convencional, ou, analisando por esta perspectiva novamente as palavras do Dr. Lavoué, ficamos até perplexos com declarações e confissões como que implorando soluções, ou denunciando a falta delas, como: “Estudar o conservadorismo morfológico a longo prazo é difícil nos sistemas contemporâneos, porque poucas linhagens animais existentes são conhecidas por permanecerem morfologicamente estáticas em relação às escalas de tempo geológicas”. (Lavoué et al, 2011). (Grifo nosso)
Desde o escândalo propalado pelo pontualismo de Eldredge e Gould, que parece que a academia representada por evolucionistas mais apegados tenta justificar ao máximo o sistema vigente geocronológico, que vai sendo solapado aos poucos por questionamentos cada vez mais irônicos, beirando a contundência e cada vez mais abertos, sem romper totalmente com o hímen do paradigma. Existe um grupo de publicações justificadoras da PMS que apelam, entre outras maneiras justificadoras, para “seleção estabilizadora”, ao mesmo tempo que acumulam outras soluções confessando indiretamente ou às vezes diretamente (Voge, 2016) sua insuficiência, pois apenas uma grande catástrofe como a queda de um grande bólido, os quais geram grandes taxas sedimentares (Sadler, 1981) e teríamos que ter morfologias não estabilizadas junto com estabilizadas, exigindo extraordinária sorte para aqueles que defendem essa tese, para que a mesma catástrofe não sepultasse as não estabilizadas pela seleção natural, exceto se a seleção natural fosse agir em conjunto com todas as espécies para formar padrão de estabilidade. Observe que a seleção, que deveria estar selecionando pelo menos alguns "díspares" para que se dirigissem no sentido de formar a árvore evolutiva de ascendência totalmente comum no registro fóssil , está agora, diante da PMS , destacando e dando maior notoriedade , dentro de diversas características contingenciais da seleção natural, sua característica conservadorista de estabilizar formas (conservadorismo que em outras circunstâncias e questionamentos é até evitado) . Os argumentos e estratagemas de destacar pontos verdadeiros, mas as vezes de forma isolada, se multiplicam em modelos computacionais, taxas flutuantes, e a confissão repetida de que tais justificativas ainda são insuficientes (Ver Tabela 2).
Tabela 2
|
Fato Surpresa |
Lista de algumas publicações justificando as anomalias não previstas pela Teoria |
|
PMS Paradoxo da Estase Morfológica
|
1. Van Bocxlaer, Bert; Hunt, Gene (20 de agosto de 2013). «Morphological stasis in an ongoing gastropod radiation from Lake Malawi». Proceedings of the National Academy of Sciences of the United States of America. 110 (34): 13892–13897. ISSN 0027-8424. PMID 23924610. doi:10.1073/pnas.1308588110 2. Lavoué, Sébastien; Miya, Masaki; Arnegard, Matthew E.; McIntyre, Peter B.; Mamonekene, Victor; Nishida, Mutsumi (7 de abril de 2011). «Remarkable morphological stasis in an extant vertebrate despite tens of millions of years of divergence». Proceedings of the Royal Society B: Biological Sciences. 278 (1708): 1003–1008. ISSN 0962-8452. PMID 20880884 3. Pigliucci, Massimo (1 de junho de 2009). «An Extended Synthesis for Evolutionary Biology». Annals of the New York Academy of Sciences (em inglês). 1168 (1): 218–228. ISSN 1749-6632.doi:10.1111/j.1749-6632.2009.04578.x 4. Estes, Suzanne; Arnold, Stevan J. (1 de fevereiro de 2007). «Resolving the Paradox of Stasis: Models with Stabilizing Selection Explain Evolutionary Divergence on All Timescales.». The American Naturalist. 169 (2): 227–244. ISSN 0003-0147. doi:10.1086/510633. Disponível em 12/09/2017 em http://www.journals.uchicago.edu/doi/10.1086/510633 5. Hunt, Gene; Hopkins, Melanie J.; Lidgard, Scott (21 de abril de 2015). «Simple versus complex models of trait evolution and stasis as a response to environmental change». Proceedings of the National Academy of Sciences of the United States of America. 112 (16): 4885–4890. ISSN 1091-6490. PMID 25901309. doi:10.1073/pnas.1403662111 6. Voje KL.Evolução . 2016 Dez; 70 (12): 2678-2689. doi: 10.1111 / evo.13090. Epub 2016 9 de novembro. 7. Alexis Matamoro-Vidal , Charlotte Prieu , Carol A. Furness , Béatrice Albert , Pierre-Henri Gouyon , Estase evolutiva na morfogênese do pólen devido à seleção natural, New Phytologist , 2016 , 209 , 1, 376 8. Monique Nouailhetas Simon , Fabio Andrade Machado , Gabriel Marroig , altos constrangimentos evolutivos limitados respostas adaptativas a mudanças climáticas passadas em calaveras de sapo, Procedimentos da Sociedade Real B: Ciências Biológicas , 2016 , 283 , 1841, 20161783 9. Florian C. Boucher, Vincent Démery, Inferring Bounded Evolution in Phenotypic Characters from Phylogenetic Comparative Data, Systematic Biology, 2016, 65, 4, 651 10. Davis, Charles C.; Schaefer, Hanno; Xi, Zhenxiang; Baum, David A.; Donoghue, Michael J.; Harmon, Luke J. (22 de abril de 2014). «Long-term morphological stasis maintained by a plant-pollinator mutualism». Proceedings of the National Academy of Sciences of the United States of America. 11 (16): 5914–5919. ISSN 1091-6490. PMID 24706921. doi:10.1073/pnas.1403157111 |
|
Tecidos moles não resistentes e não fossilizados (permineralizados) |
1. Butterfield NJ Conservação orgânica de organismos não mineralizantes e a torneira do Burgess Shale. Paleobiologia. 1990; 16 : 272-286. 2. Stankiewicz BA, Briggs DEG, Michels R, Collinson ME, Flannery MB, Evershed RP Origem alternativa do polímero alifático no querogênio. Geologia. 2000; 28 : 559-5 3. Nguyen RT, Harvey HR Preservação de proteínas em sistemas marinhos: associações hidrofóbicas e outras não covalentes como principais forças estabilizadoras. Geochim. e Cosmochim. Acta. 2001; 65 : 1467-1480. 4. Briggs DEG O papel da decadência e da mineralização na preservação de fósseis de corpo macio. Ann. Rev. Earth Planet Sci. 2003; 31 : 275-301. Excepcional preservação fóssil e a explosão cambriana. 5. Butterfield NJ Integr Comp Biol. 2003 fev; 43 (1): 166-77. 6. Schweitzer, Mary Higby; Wittmeyer, Jennifer L; Horner, John R (22 de janeiro de 2007). «Soft tissue and cellular preservation in vertebrate skeletal elements from the Cretaceous to the present». Proceedings of the Royal Society B: Biological Sciences. 274 (1607): 183–197. ISSN 0962-8452. PMID 17148248. doi:10.1098/rspb.2006.3705 |
|
Presença de quantidade datável de Carbono 14 em Fósseis e minerais de Ma |
1. "Use of natural diamonds to monitor C-14 AMS instrument backgrounds," Nuclear Instruments and Methods in Physics Research B 259, (2007) p. 282-287. 2. Quaternary geology, radiocarbon datings, and the age of australites," Geol. Soc. America Spec. Paper, 84, (1965) p.415-432. 3. Dating Infall of Australites," (1970), Radiocarbon Journal 13(1), (1970) p. 8-11 4. Holzschuh, Josef (2005). «Recent C-14 Dating of Fossils including Dinosaur Bone Collagen». scienceeevolution. Consultado em 9 de setembro de 2017 |
|
Inúmeros artigos denunciando anacronismos |
180 artigos confrontando datação radiométrica «Search Tools | The Institute for Creation Research». www.icr.org (em inglês). Consultado em 14 de setembro de 2017
|
Observe que a seleção, que outrora precisaria selecionar "díspares" para que se dirigissem no sentido de formar a árvore evolutiva de ascendência totalmente comum no registro fóssil, está agora , como um coringa, diante do PMS, sendo lida e dando destaque a seu poder conservadorista de estabilizar formas. Na revisão bibliográfica de Voge assim se resume:
"A afirmação de que a mudança direcional raramente foi observada no registro fóssil e que a estase era o modo dominante nas linhagens evolutivas (Eldredge e Gould, 1972, Gould e Eldredge 1977, Cheetham 1987, Jackson e Cheetham, 1999) provocaram um intenso debate entre biólogos evolutivos (Gould 1980, Charlesworth et al., 1982), que ainda está em andamento (por exemplo, Lieberman e Eldredge 2014, Pennell et al., 2014a, b, Venditti e Pagel 2014) Parte do legado da teoria do equilíbrio pontuado (Eldredge e Gould, 1972; Gould e Eldredge 1977; Gould 2002) é a constatação de que as espécies estabelecidas geralmente mostram uma evolução líquida mínima ao longo do tempo no registro fóssil. Também se verificou que as diferenças entre as espécies no registro fóssil são menores do que o esperado devido à deriva genética sozinha (p. Ex., Lande 1976; Lynch 1990; Cheetham et al. 1993). O domínio da evolução lenta no registro fóssil não é diretamente derivado do que sabemos dos estudos microevolutivos: os traços quantitativos geralmente apresentam variação genética substancial (Houle 1992, 1998; Hansen et al., 2011), uma seleção forte é comum (Hereford et al. 2004), e uma quantidade considerável de evolução é freqüentemente observada em intervalos de tempo de anos a algumas décadas (Hendry e Kinnison 1999, Kinnison e Hendry 2001, Hendry et al., 2008, mas veja Merila et al., 2001). As observações contrastantes da mudança de evolução em escalas de longo e curto tempo são conhecidas como o "paradoxo da estase" (Wake et al. 1983; Hansen e Houle 2004; Futuyma 2010) e faz a história de longo prazo de A vida parece quase desacoplada do processo evolutivo que estudamos em escalas de tempo mais curtas. A afirmação de que a mudança direcional raramente foi observada no registro fóssil e que a estase era o modo dominante nas linhagens evolutivas (Eldredge e Gould, 1972, Gould e Eldredge 1977, Cheetham 1987, Jackson e Cheetham, 1999) provocaram um intenso debate entre biólogos evolutivos (Gould 1980, Charlesworth et al., 1982), que ainda está em andamento (por exemplo, Lieberman e Eldredge 2014, Pennell et al., 2014a, b, Venditti e Pagel 2014) "
Tabela 2
O presente estudo vai isentar a causa dessas explicações ad hoc: Que julgamos ser a própria submissão e o não questionamento ao uniformitarianismo da constância de decaimento radioativo, que nos “assegura” que como ratos presos na ratoeira, só nos resta comer o queijo e justificar porque a evolução quase parou em meio bilhão de anos (isso se considerarmos apenas os fósseis do fanerozóico, pois se formos falar de bactérias e fósseis pré-cambrianos teríamos que assumir estase morfológica em 3,5 bilhões de anos). Ou como podemos observar nesta tabela que representa apenas uma pequena amostra desta dependencia:
Padrões Fósseis Contrastando Ausência e Presença de Evidências
|
Fatos |
Evidência de ausência |
Evidência de presença |
|
Paradoxo da estase morfológica PMS |
De comportamento plástico, geneticamente dinâmico até de espécies com os mesmos genótipos, que estariam em evolução durante milhões de anos, intercalados por catástrofes e intensas mudanças ambientais. |
De um mesmo ambiente em todo o planeta, sistema homogêneo, ou segregado e bastante adaptável as formas de vida, poderia gerar estase morfológica. |
|
Pobreza taxonômica (250 a 300 mil espécies) sobre estimativas de haver inumerável amostras fósseis. |
De especiação incluindo e sobretudo em tempo real, diversidade, o que resultaria em riqueza de diversificação fóssil e menor disparidade de padrões, pela presença de maior número de padrões morfológicos. A pobreza taxonômica revela ausência de motores evolutivos atuando, o que implica em ausência de tempo geocronológico, que caso houvesse, permitiria a evolução como fato observado, ser identificada na evolução histórica. |
O modelo de Sepultamento de populações ancestrais comuns dos quais derivariam a biodiversidade atual, exceto das morfologias extintas, se revela extremamente coerente com este padrão fóssil. |
|
Repetição de mesmas espécies e permanência de 4229 gêneros denominados fósseis vivos (Romer, Whitimore) |
De previsão de que as espécies mudam, se especiam, se transformam, não ignoram mudanças ambientais. |
Sepultamento de populações repetindo mesmas espécies no registro fóssil, recomendando modelo de segregação e estratificação espontânea como melhor inferência para a estratificação fóssil, bem como observações ligadas a flutuabilidade de corpos. |
|
Padrão de surgimento repentino de arquétipos prontos, de planos corporais e sistemas de vida prontos |
De saltacionismo ou gradualismo que deveria haver no registro fóssil, anterior ao surgimento destes arquétipos e planos morfológicos, corporais e sistemas de órgãos e células. |
De que a paleontologia se comunica com a idéia de necessidade de surgimento pronto de sistemas com complexidade irredutível. |
|
Em bivalves (conchas do mar) houve desaparecimento de morfologias (a nível de famílias) aumentando desde o cambriano, chega a um clímax no permiano e vai caindo a extinção lentamente até as camadas recentes, formando uma parabólica |
Nenhuma previsão para ordenar o aumento de extinção, chegando ao ponto máximo e clímax justamente no momento mais dramático da terra (permiano), para depois haver igual lenta diminuição. Ausência de previsão de que as catástrofes fossem aumentando e depois diminuindo.
|
Um momento no planeta repleto de Catástrofes se iniciando, decantando nas camadas mais profundas, aumentando suas deposições erodidas e extinguindo e sepultando vivos cada vez mais, chegando a um clímax e depois diminuindo seus rastros sedimentares fossilizadores, diminuindo cada vez mais a extinção dos que sobreviveram cada vez que as catástrofes diminuísse suas transgressões e regressões marinhas. |
|
Desaparecimentos coordenados nos topos de estratos . “Dinossauros e muitos outros grupos de répteis e invertebrados marinhos desaparecem do registro no topo do Mesozóico. Outros exemplos de desaparecimentos coordenados em larga escala ocorrem na parte superior do ordoviciano, perto do topo do Devoniano e do topo do Triássico.” |
De razão para tal desaparecimento. |
Segregação e estratificação espontânea (Berthaut, 2008. Maske, 2010) explica a separação de tipos sedimentos em ajuntamentos comuns e pode explicar o desaparecimento em determinadas camadas geológicas ou posições mais altas das mesmas, já que a tendência de corpos é afundar. |
|
A diminuição da diversidade de microfósseis nas rochas pré-cambrianas. (Schopf JW, 1992) |
De comportamento característico de bactérias de povoarem indeterminadamente quando estão sem competidores |
O processo de lixiviação e filtragem para camadas estando embaixo das sedimentares ou estando em altitudes as quais recebem menor infiltração contendo futuros microfósseis sob águas contendo sílica o suficiente para o processo de fossilização. |
Definições de Espécie, gênero e famílias, e MPTG
Durante décadas, a biologia evolutiva, diante da “plasticidade dos seres vivos” muito citada desde Spencer, 1820-1903 (Lightman & Bernard, 2016), e tão destacada sobretudo desde a imensa revisão bibliográfica feita, quando ainda jovem, por Charles Darwin, tem dificuldades para entender e definir o significado de “espécies”. Atualmente, existem mais de vinte e dois conceitos diferentes de espécies (de Queiroz, 2005). O uso de diferentes conceitos leva a comparações desencontradas na ciência que visa por sistematizar e organizar o conhecimento. Por outro lado, biólogos catastrofistas por décadas têm utilizado para espécies dinâmicas e plásticas, termos e conceitos apropriados mais abrangentes e flexíveis como “tipo” ou “grupo” para o que consideram categorias de organismos relacionados geneticamente, que, em testes de cruzamentos artificiais, gerem emparelhamento cromossômico e pelo menos o início de embrião com características paternas e maternas (Junker & Scherer, 1996). Assim, torna-se prático o agrupamento de clados em torno destes testes de compatibilidade no crossover.
Cada uma das várias categorias de espécies, subespécies e variedades que vemos hoje foram concebidas para se diversificarem de um tipo básico ancestral comum fóssil, portanto consideraremos neste trabalho, a grandeza MPTG (Morphological patterns around the taxon genus), padrões morfológicos em torno de gêneros . Esta grandeza se identifica com fósseis ancestrais, com a falseável hipótese dos tipos básicos (Marsh, 1941), já que “as novas formas tornam-se cada vez mais refinamentos das existentes formas” (Benton et al, 2007). Entre outras observações , podemos dizer que a disparidade morfológica no registro fóssil, quando comparamos táxons mais altos, é alta , e a diversidade de espécies, no registro fóssil, é baixa. Ou seja, um verdadeiro espelho de ancestrais básicos dos quais a maioria de nós (enquanto biodiversidade) descendemos. Analogamente, Muitos troncos e Arbustos com galhos grossos (disparidade), no registro fóssil versus arbustos com infinitas folhagens e raminhos na atualidade(diversidade) . Isso não anula variações morfológicas no registro fóssil , claro que elas existem dentro de um morfoespaço que abriga inclusive mesmos genotipos, e é até esperado que existam evolutivamente , pois as espécies mães possuem cada vez mais , maior pool gênico, uma vez que ainda não estavam desgastadas pelas sucessivas derivas e motores adaptacionais , stressantes e altamente empobrecedores (Mayr), nem pelo acúmulo de genes deleterios que nos lega o motor da entropia genética (Sanford, 2005, 2017; Crabtree, 2010).
Diversos exemplos de disparidade e diversidade são citados na literatura (Benedict et al, 2016). Ou seja, defendemos que MPTG ancestrais básicos em suas características de disparidades morfológicas (e baixa diversidade ) foram sepultados, em geral catastroficamente e com “morte súbita”, para se tornarem fósseis, e este modelo nos conta uma história de um período de um momento catastrófico geológico de: 1) Falta de especiação rápida ou lenta, o que, caso houvesse, enriqueceria a taxonomia fóssil, assim como temos hoje na biodiversidade atual (Benton et al, 2007; Futuyama, 2010) 2) Permanência com alto número de mesmas espécies, o que necessitaria de ambiente estável (repetição fóssil sem pressões ambientais que provocariam manifestações adaptativas e evolutivas), 3) Desastre soterrador de população viva evidenciada pelas repetições de mesmas espécies fósseis (o que descaracteriza a pontualidade, tempo longo, e substituiria a parcial sucessão faunística por por “segregação e estratificação espontânea” SEE (Minoletti, 2009, Berthaut, 1986, 1988) que explicam a presença de diversas espécies diferentes segregadas no registro fóssil, bem como não precisam apelar para "ad hoc" para explicarem árvores polistratos e milhares de anacronismos 5) Fósseis de imensos vertebrados completos (o que caracteriza desastre de grande magnitude global e altas taxas de sedimentação, sobretudo de vertebrados completos terrestres), e 6) Mudança drástica no ambiente gerando a radiação adaptativa das espécies nas camadas recentes nas milhões de espécies da biodiversidade atual diversificada por mais que mantenha semelhante numero de padrões corpóreos (Wise, 2013).
Muitas das espécies e suas variações, observáveis hoje, refletem os mesmos padrões morfológicos em torno do táxon gênero (MPTG), apesar de grande parte da literatura designar o táxon superior famílias como padrões morfológicos, o que resulta no fato de lermos na literatura "extinção de famílias "( o que na verdade pode ser apenas gêneros) . Sobre este problema, depois de consultar diversos especialistas e literatura solicitando dados globais sobre quantidade de especies , generos e familias em cada periodo geológico, e receber respostas lacônicas sem solução , comçamos a entender declarações como: “Atualmente, não há uma definição acordada de disparidade, muito menos qualquer consenso sobre como medir” (Wills et al, 1994), então escolhi como melhor síntese a resposta por e-mail, em 2013, encaminhada pelo paleontólogo de Harvard , orientado por Stephen Jay Gould, Dr. Kurt Wise, o qual praticamente repete (porém com maior riqueza) a mesma observação de especialistas consultados, de que as definições e organizações, devido haver inúmeros critérios, estão em falta:
“The best early estimates (by Dave Raup and Jack Sepkoski in the 1980s) were based upon a family-level fossil record data (namely, Sepkoski’s family-level compilation of the marine fossil record). From the number of families that went extinct, the current intra-family species diversity was assumed to be true of the fossil families, and the necessary % species extinction was calculated that would result in that much family extinction (by computer bootstrapping, etc.). Later, after Jack Sepkoski had compiled genus-level data for the marine fossil record (2002, Bulletins of American Paleontology 363), the same sort of calculations were done with the genera that had earlier been done with families. Not only have the species-level data not yet been compiled, but many paleontologists consider that data would be unreliable (because of differences of opinion on how to define paleontological species)”.
Sendo a classificação a nível de famílias algo muito contraditório quando entendemos que família agrupam gêneros que não foram extintos, é comum lermos na literatura dados referentes a uma coisa e outra ao mesmo tempo, ou sobre percentuais ocorridos nas extinção em massa sem o número anterior de onde se retirou tais percentuais. Por mais que isso não tenha gerado tantas correções a nível de descrever extinções (interpretação) ou diminuição de fósseis na estratigrafia (fato), (Sepkoski, 1993), isso inviabiliza ou dificulta outras percepções sobretudo de relação evolutiva no estudo paleontológico. Recentemente gêneros não extintos de bivalves foram classificados como famílias (Gibson, 1996). E devido a isso, tentarmos entender MPTG como uma maneira flexível de lidar com estas imprecisões, o que pode nos ajudar na organização do entendimento geral destas dificuldades classificatórias.
Portanto, MPTG, é o que consideramos como as características peculiares que vão desde os dos tipos básicos matrizes fósseis , até sua descendente diversificação no presente, que ainda hoje estão, exceto famílias e “gêneros” extintos, presentes na maioria das diversas espécies e suas variações encontradas na natureza (Eldredge e Stanley, 1984). Muitas espécies viventes hoje, e suas variações, refletem padrões morfológicos semelhantes. A permanência destes mesmos padrões morfológicos fósseis, por mais que aumente o número de espécies, facilita percebermos os limites da evolução em torno de MPTG , situando e orientando as mudanças possíveis de serem registradas no registro fóssil e testadas seus parentescos na atualidade (Junker & Scherer, 1996). Também o fato desta descontinuidade estar ainda mais marcante no registro fóssil, quanto mais superiores táxons, fortalece esta idéia de descontinuidade sistemática (Carroll, R. L. ,1992).
A tese dos tipos básicos matrizes ou ancestrais (Marsh, 1941), tem sido apoiada pelo fato de o registro fóssil apresentar baixa variabilidade (estase morfológica) e baixa radiação especiativa entre as espécies fósseis (Zimmerman, 1960; Martens, 1997; Albrecht e Wilke, 2008; Alisson, 2013), independente da suas respectivas plasticidades ou maleabilidades fenotípicas ou genotípicas (Ghalambor et al., 2015). Além disso, outros fatores que corroboram a tese dos tipos básicos ancestrais são os mais de 4229 gêneros bem documentados de “fósseis vivos” que são chamados assim por terem sofrido poucas mudanças ao longo do tempo, logo, mantendo-se semelhantes às encontradas no registro fóssil (Romer, 1966; Whitmore, 2013a; Whitmore, 2013b). Provavelmente refletindo idéias de sua época, Darwin chegou a citar essa dedução corrente ao escrever:
"Não há uma verdadeira grandeza nesta forma de considerar a vida, com os seus poderes diversos atribuídos primitivamente pelo Criador a um pequeno número de formas, ou mesmo a uma só? Ora, enquanto que o nosso planeta, obedecendo à lei fixa da gravitação, continua a girar na sua órbita, uma quantidade infinita de belas e admiráveis formas, saídas de um começo tão simples, não têm cessado de se desenvolver e desenvolvem-se ainda!... O venerável e reverendo W. Herbert, mais tarde deão de Manchester, escrevia em 1822, no 4., volume das Horticultural Transactions, e na sua obra as Amaryllidacées (1837, p. 19, 339), que ‘as experiências de horticultura têm estabelecido, sem refutação possível, que as espécies botânicas não são mais que uma classe superior de variedades mais permanentes’. Aplica a mesma opinião aos animais e vê que as espécies únicas de cada gênero foram criadas num estado primitivo muito plástico, e que estes tipos produziram ulteriormente, principalmente pelo cruzamento e também por variação, todas as nossas espécies existentes. Na Nouvelle Flore de l’Amérique Du Nord (1836, p. 6), Rafinesque exprimia-se assim: ‘Todas as espécies podiam ser outrora variedades, e muitas variedades tornaram-se gradualmente espécies, adquirindo caracteres permanentes e particulares’; e um pouco mais adiante (pág. 18) acrescenta: ‘excetuando os tipos primitivos ou ancestrais do gênero’”(Darwin, 1866, capítulo 9, pp.25-26, 577, ênfase nossa).
Quando falamos de estase morfológica no registro fóssil, nos referimos às camadas sedimentares compreendidas entre o ediacara/cambriano até próximo das camadas do período Pleistoceno, ou camadas superficiais, como demonstra diversas publicações baseadas na teoria do equilíbrio pontuado (pontualismo) proposta pelos paleontólogos Niles Eldredge e Stephen Gould (Levinton e Chris, 1980; Woodruff, 1980; Williamson, 1981; Eldredge, 1986; Van Bocxlaer e Hunt, 2013). E não nas camadas atuais onde a variação irradia em multiformes variações e ocorre fossilização também.
Os padrões morfológicos ligados ao táxon gênero MPTG, podem ser observados no registro fóssil surgindo repentinamente em todas as camadas sedimentares, com expressivo aparecimento no cambriano e surgimentos abruptos sem nenhum gradualismo em toda coluna geológica, mas uma repetição surpreendente. Essa constatação tem sido divulgada há décadas pelos adeptos do pontualismo que ainda admitem a estase morfológica e a radiação florescendo em especiações somente nas camadas geológicas mais atuais (Levinton e Chris, 1980; Woodruff, 1980; Williamson, 1981; Eldredge, 1986; Van Bocxlaer e Hunt, 2013). Estudo evolutivo recente corrobora tal afirmação ao dizer que :
"A visão dominante da evolução baseada no registro fóssil é que as espécies estabelecidas permanecem mais ou menos inalteradas durante sua existência. Por outro lado, a evolução substancial é rotineiramente relatada em populações contemporâneas, e a maioria dos traços quantitativos mostra um alto potencial de evolução. Essas observações contrastantes em escalas de tempo longo e curto são muitas vezes referidas como o paradoxo da estase, que se baseia na suposição fundamental de que os períodos de estase morfológica no registro fóssil representam uma mudança evolutiva mínima." (Voje, 2016).
Assim, o presente artigo tem como objeto de estudo esses fatos observados e bem documentados: surgimento repentino fóssil, estase morfológica, repetição de mesmas espécies fósseis em contraste com a explosão de radiação adaptativa revelando todo potencial de plasticidade e maleabilidade dos fenótipos e genótipos dos seres vivos que deveriam estar presente no registro fóssil caso a história evolutiva tivesse ocorrido ali, pois “a diversidade morfológica diminui juntamente com a diversidade taxonômica. Este padrão sugere heterogeneidades como a extinção elevada e / ou a origem reduzida em certas regiões do morfoespaço” (Foote, 1993).
As especiações e variações que, como vimos, carregam os MPTG podem ser observadas ocorrendo em tempo real e histórico-arqueológico, mas estão ausentes na mesma proporção, no registro fóssil. Isto se torna relevante na medida em que percebemos o sucesso reprodutivo das variações e sua permanência ao longo do tempo, pois a variação em torno de gênero não exige grandes bio-transformações, mas deve ocorrer de qualquer forma, em torno do gênero (MPTG) nas amostras fósseis. Hoje, surpreendentemente, segundo o paleontólogo especialista em estatísticas fósseis Dr. Kurt Wise, tecnicamente, o número de planos corporais no registro fóssil não excede o número de planos corporais hoje (Wise KP. 1989), e segundo esta afirmação, temos quase o mesmo número de padrões corporais entre os 250-300.000 fósseis catalogados, comparados aos 2 milhões de espécies vivas hoje já catalogadas, com estimativas de 8,7 milhões e estimativas que vão até 100 milhões de espécies hoje) e as ínfimas 300 mil espécies fósseis do cambriano/edicara ao pleistosceno. (Woodmorappe, 2000; Sadava et al., 2009; Mora et al., 2011; Catalogue of Life, 2016). Quando comparados os padrões morfológicos, esperar-se-ia que tivéssemos um número bem maior de PMTG no registro fóssil se este espelhasse uma amostra da biodiversidade ocorrida em 544 milhões de anos correspondentes ao Fanerozóico. Portanto, o fato do número de espécies atuais, na casa dos milhões, não conseguir transpor o número de padrões corporais presente no registro fóssil, sugere evolução delimitada a padrões morfológicos ou tipos básicos ancestrais (baraminologia) ou MPTG.
Especiação em Tempo Real
A surpresa de muitos diante da especiação rápida e mudanças significativas morfológicas em apenas uma geração, refletia a falta de previsão da teoria do que se esperaria em termos de tempo.
Entendemos a especiação em tempo real como um fenômeno em que duas ou mais populações de uma mesma espécie, se modificam em novos arranjos de informação genética pré-existente, tendo como causa a separação por barreiras geográficas e mortandades, porém em um tempo hábil para que seja possível a observação de todo o processo do início ao fim. (Furness et al 2015). Experiências empíricas promovendo especiação em tempo real , tem sido testadas com sucesso e colocando em xeque cada vez mais o PMS, (Ghalambor et al, 2015), o que demonstra o que “nenhum organismo ignora seu ambiente” (Ezard et e al, 2016).
Podemos ainda classificar o processo de especiação de duas formas: em tempo real ou em tempo histórico-arqueológico. A especiação em tempo real é aquela em que ocorre bio-modificações limitadas ao tipo básico, equivalentes ao surgimento rápido de uma nova “espécie”, observada ̶ por experimentação ou observação não-controlada ̶ em até 50 anos. A especiação em tempo histórico, por sua vez, diz respeito às bio-modificações equivalentes a uma nova “espécie” acima de 50 anos que podem ou não ser observadas devido o tempo de vida do observador ou projeto de pesquisa. Nesses casos, há 50% de chance de observação e os outros 50% restantes dependem do uso da dedução, por exemplo, a partir de análise cromossômica e cálculos de taxas de radiação (Trivedi, 2000).
O papel da deriva genética no processo de especiação
Existem diversos mecanismos envolvidos no processo de especiação (epigenética, deriva genética, seleção natural, influências ambientais e geomagnéticas no crossover (Gorelick, 2005), etc). Devido à especiação estar intimamente relacionada à deriva genética e a conseqüente perda de pool gênico de uma população anterior, estudos do modelo de muitos ancestrais básicos (SRABUC) que apostam na especiação em tempo real, se tornam fundamentais para explicar as evidências atuais de limites à adaptação nos diferentes organismos vivos (Bell, 2013).
A deriva genética é um mecanismo que modifica aleatoriamente e de forma repentina – devido a catástrofes ou a isolamentos diversos, por exemplo, − as frequências alélicas de uma população (Ridley, 2006; Freeman e Herron, 2009). Por outro lado, acredita-se que a seleção natural é direcionada, isto é, elimina muitas mutações deletérias, mutações necessárias se perdem juntas, ignora as mutações neutras e deletérias (principalmente as que se manifestam após período reprodutivo) seleciona os traços sobreviventes e/ou vantajosos pré-existentes ou que comunicaram em face de resposta epigenética ao meio (Sanford, 2014). Porém, como afirmou a bióloga americana Lynn Margulis, “a seleção natural elimina, e talvez mantenha, mas ela não cria” (Teresi, 2011, p. 68). Ademais, devido o fato da deriva genética não fazer distinção entre genes bons e ruins, e a seleção natural falhar também em seu filtro, isso resulta em perda de variação genética (Lacy, 1987), deixando as espécies geneticamente mais pobres e “cada vez mais próximas da extinção”. Em outras palavras, a deriva decepa e empobrece a população de seu “pool gênico” e a seleção natural ajeita mais ou menos o que restou.
Nesta frase farei uma modificação, pra que fique mais técnico.Sabe-se que o efeito da deriva é inversamente proporcional ao tamanho da população, podendo aparecer em diferentes momentos da história das espécies e da própria humanidade. Nesse sentido, é possível que a deriva tenha tido um papel fundamental e sobrepujante ao da seleção natural no que diz respeito ao processo de especiação rápida após uma grande catástrofe.
O processo de especiação após Catástrofes
Grandes catástrofes mundiais são as formadoras da maioria das camadas sedimentares presentes no globo, que estão associadas a imensas inundações e destruições em todo globo (Souza Jr, 2008). As conseqüências da mortalidade em massa gerada, stress endogâmico nas espécies sobreviventes e isolamentos geográficos, deram, alem de outros fatores, condições muito favoráveis para uma rápida especiação (Wilmer et al., 2011). O modelo SRABVSURC defende o sepultamento da biodiversidade de todo planeta por estratificação espontânea nas transgressões e regressões marinhas (Berthault, 1986; Berthault, 1988; Brand e Tang, 1991;Snelling, 1997; Berthault, 1998; Chadwick e Spencer, 2006; Berthault, 2013), seguido de especiações rápidas pelos motores evolutivos adaptacionais que diversificariam os padrões de ancestrais prontos sepultados em um catastrofismo inédito (SRABURC- Standard of Ready Ancestors Buried in Unprecedented Catastrophism ).
O modelo SRABVSURC de história curta da formação das camadas sedimentares prevê um momento em que os poucos seres humanos e animais sobreviventes e isolados por essas catástrofes associadas (de energia e magnitude global) teriam recomeçado a repovoar a terra, e sucessivas derivas e stress endogâmicos de cruzamentos consangüíneos haveriam de ocorrer, exatamente como percebemos em espécies de Denisovans (Prüfer et al., 2013) e nas fundações de etnias de biotipos semelhantes entre si habitando em cima de fósseis de halogrupos dessemelhantes como, por exemplo, mongolóides nas Américas em cima de fósseis dessemelhantes negróides em Lagoa Santa-MG, São Paulo e México. Fato observado também na China de mongolóides habitando em cima de fósseis caucasianos e negroides (Ke, Y, 2001) tanto nas etnias humanas quanto em milhares de outros seres vivos semelhantes entre si isolados de seus parentes.
...” negróides e australóides, voltando a levantar uma grande polêmica que coloca em questão os modelos de povoamento pré-histórico da América. Reforçando a possibilidade de ser a população de Lagoa Santa, como outras populações americanas de mais de 7 mil anos, proveniente de migração muito antiga de grupos não mongolizados, estes novos achados trouxeram de volta e com força para o cenário científico, uma polêmica de 160 anos.”( Mendonça de Souza, 2006)
Nesse contexto, a especiação em tempo real legitima o modelo de um “salto evolutivo” recente, com dois picos bio-diferenciadores relacionados aos episódios do inicio da entropia genética humana (Fu et al., 2013) depois das catástrofes associadas que empobreceriam o pool gênico dos seres vivos e dariam aos sobreviventes, em virtude da migração e do isolamento geográfico, o legado de efeito fundador – situação frequente na especiação peripátrica− em um planeta com um ecossistema totalmente reconfigurado onde o motor epigenético atuaria para corresponder as necessidades do novo ambiente (Ridley, 2006; Eakin, 2014; Weyrich et al., 2016).
A especiação peripátrica é um mecanismo pelo qual podemos explicar o enorme aumento da diversificação pós-catástrofes. É um tipo de especiação pelo qual as espécies novas são formadas em populações periféricas isoladas (Ridley, 2006). Na especiação peripátrica, populações drasticamente reduzidas fazem com que a especiação completa seja o resultado mais provável do isolamento geográfico, porque a deriva genética age mais rapidamente em populações pequenas. Deriva genética somada a fortes pressões seletivas, causariam uma rápida mudança genética na pequena população descendente (Wilmer et al., 2011).
Observações atuais podem servir de exemplo para compreendermos melhor os casos de especiação peripátrica. Estudo recente afirma que “uma análise de mais de 2.000 espécies de pássaros fornece uma visão sobre como evoluíram as diversas formas de bico dos animais e aponta para um único evento raro como um gatilho para a rápida divergência inicial das linhagens aviárias.” (Bhullar, 2017).
Toda essa sucessão de fatos pode ser vislumbrada por meio de um cenário onde a maioria dos organismos destruídos por essas grandes catástrofes associadas e consequentes, deixaria pequenas populações de sobreviventes (Wilmer et al., 2011). Segundo matéria publicada pela Folha de São Paulo a respeito deste achado, “o que pareceu fascinar mais outros biólogos, porém, é a grande velocidade com que o fenômeno do deslocamento de caráter ocorreu. ‘Eu acreditava que fosse demorar muito mais’, comentou na ‘Science’ o biólogo David Pfening, da Universidade da Carolina do Norte. A redução média de 5% no tamanho de bico, considerada drástica pelos biólogos, ocorreu no intervalo de cerca de um ano, praticamente de uma geração para a outra.” (Folha de S. Paulo, 2006).
A surpresa dos biólogos evolutivos com a descoberta desse novo fenômeno (Grant, 2017) se dá devido à falta de entendimento de outros motores coadjuvantes, que atuam nessas bio-modificações limitadas ao tipo básico, tais como epigenética e aspectos energéticos e de temperatura atuando no crossover (Fondon e Garner, 2004; Eakin, 2014). Para a Dra. Jean K. Lightner,45 parece haver três fontes para as variações associadas a radiação adaptativa: 1) hibridização, 2) mutação e 3) triagem ambiental de alelos ancestrais (por mecanismos de seleção natural e deriva meiótica) (Lightner, 2016).
Estas observações, se não encarecem a dívida dos elos perdidos na paleontologia pontualista, aumenta muito a dívida de variabilização taxonômica fóssil para o modelo histórico geológico que assume pelo menos cinco grandes catástrofes separadas por milhões de anos na história da terra.O curioso é que as pesquisas ligadas a especiação em tempo real tanto confirmam as observações de Darwin no âmbito da biologia funcional (equilíbrio pontuado) quanto destrói por completo os postulados evolutivos em termos de períodos geológicos (gradualismo filético), e é perfeitamente comunicável a modelos catastrofistas que aposta numa especiação rápida impulsionada pelo efeito das bio-modificações limitadas ao tipo básico.
Baraminologia e o estudo dos “tipos básicos”
O aparecimento repentino e pronto de formas de vida e de sistemas irredutivelmente complexos (Lonning, 2005), além do número tecnicamente igual (Wise, 1989) de padrões morfológicos quando comparamos taxonomia fóssil de 250-300 mil espécies com estimativas de 9 a 100 milhões atuais, sugere a idéia de um fenômeno de tipos básicos ancestrais surgidos prontos e a permanência numérica de padrões corporais de uma evolução limitada em torno deles (MPTG). Há uma comunicação muito fluente entre o darwinismo biológico e o modelo SRABVSURCbiológico, onde muitos se valem de descobertas evolutivas, como especiação em tempo real, para se aproximar mais da historicidade de curto período de tempo, e utilizando a radiação recente como justificativa para toda a biodiversidade. A teoria da evolução sintética, mais especificamente atuando no âmbito biológico, que ensina a diferenciação das espécies, neste contexto, tem baixa discórdia entre as classes mais avançadas de SRABUC, sobretudo nos cientistas do movimento de “Baraminologia” (Marsh, 1941; Wise, 1992; Robinson e Cavanaugh, 1998; Frair, 2000; Jerlström, 2000; Wood e Cavanaugh, 2001; Cavanaugh e Wood, 2002; Wood, 2010; Aaron, 2014). Suas discordâncias se concentram mais no âmbito da história geo-paleontológica compreendida entre o cambriano/edicara e pleistoceno e/ou camadas superficiais.
Desde 1941, o modelo SRABVSURC é contrário a ideia de que especiação é sinônimo de “evolução” por meio de estudos relacionados às especiações no campo da Baraminologia (Marsh, 1941; Wise, 1992; Robinson e Cavanaugh, 1998; Frair, 2000; Jerlström, 2000; Wood e Cavanaugh, 2001; Cavanaugh e Wood, 2002; Wood, 2010; Aaron, 2014). O biólogo norte-americano Dr. Frank L. Marsh, um dos fundadores da Creation Research Society foi quem cunhou a palavra "baramin". (Marsh, 1941; Frair, 2000) Ela foi derivada da combinação de duas palavras hebraicas − bara (criado) e min (tipo) – referindo-se a tipos básicos criados (espécies, em versões bíblicas em português) (Frair, 2000).
Em 1990, o paleontólogo catastrofista Kurt Wise observou a necessidade de uma biossistemática SRABVSURC− um método de estudo, nomeação, e classificação de baramin (Wise, 1990; Frair, 2000) ou ancestrais de MPTG como defendemos aqui. O campo científico foi oficialmente denominado como “baraminologia”, que de forma simplista significa o estudo dos baramins ou dos tipos básicos ancestrais. Segundo os pesquisadores Reinhard Junker e Siegfried Scherer,“tipos básicos é uma unidade de classificação, um taxon, resultado do trabalho da descontinuidade sistemática como é observado na natureza” (1996, p.34; Wood et al., 2003). Dito de forma simples, tipos básicos criados variabilizaram ao longo do tempo até chegarem ao que conhecemos hoje como subespécies.
Existem algumas regras falseáveis para se considerar um grupo de espécies como pertencentes a um tipo básico ancestral comum. O Dr. Junker e o Dr. Siegrifield Scherer destacam na 6ª edição alemã do livro Evolução, um livro texto crítico:
“todos os indivíduos que estão unidos direta ou indiretamente por cruzamentos são considerados pertencentes a um tipo básico (nível genético). E todas as espécies biológicas que se se assemelham claramente umas as outras pertencem a um gênero (nível morfológico). E todas as espécies biológicas que em princípio podem cruzar entre si pertencem a um tipo básico (nível morfo-genético)”... “dois indivíduos pertencem ao mesmo tipo básico quando a embriogênese de um híbrido vai além da fase maternal do desenvolvimento e contem uma expressão coordenada e genes morfogenéticos paternos e maternos” (Junker e Scherer, 1996, p.34).
Além disso, os baraminologistas usam uma série de critérios metodológicos de adesão para determinar os limites dos grupos baramins (Robinson, 1997; Wood, 2001; Wood, 2002; Wood, 2005; Cavanaugh e Sternberg, 2004). De modo geral, os métodos mostram espacialmente graus de similaridade e de dissimilaridade entre grupos, e podem revelar informações taxonômicas úteis, distinguindo cada vez mais os fatores que dão probabilidade ou não de parentesco, aumentando assim sua contribuição em biologia aplicada a técnicas de melhoramento genético e estudo do comportamento evolutivo das populações.
A baraminologia, também conhecida como sistemática de descontinuidade, está rapidamente se tornando uma das áreas mais ativas de pesquisa SRABVSURC (Scherer, 1993), e algumas de suas metodologias têm sido aplicadas e testadas até mesmo por pesquisadores ligados ao modelo gradualista geológico de ancestralidade totalmente comum com destaques em periódicos com peer-review (Senter, 2010; Wood, 2011). Como já vimos, sua principal finalidade é determinar quais organismos compartilham um ancestral comum (Frair, 2000). A ideia básica defendida neste campo de pesquisa é a de que há limites nas possibilidades de cruzamento que não podem ser atravessados. Nesse contexto, baraminologistas objetivam encontrar as "descontinuidades" na história de vida, ou os limites da ancestralidade comum (Remine, 1993).
Esse campo de pesquisa ganha ainda mais estímulo frente às evidências atuais mostrando que “especialistas” têm erroneamente classificado algumas espécies dentro de um dado gênero por conta do “desejo” em descobrir o ancestral comum universal (Lopes, 2015). Paleontólogos afirmam que um terço das “espécies” reconhecidas como sendo de dinossauros talvez nem mesmo tenha existido (Horner e Goodwin, 2009). Para eles, essas “espécies” podem não ser espécies separadas, mas estágios juvenis ou subadultos, em desenvolvimento, identificados erroneamente como exemplares de outras espécies. Em artigo publicado na revista Science, por exemplo, Schwartz e Tattersall afirmam que esse milagre da multiplicação da nomenclatura das espécies foi longe demais (Schwartz e Tattersall, 2015).
Vale lembrar que, embora a Baraminologia tenha alcançado resultados promissores, suas conclusões não são definitivas (Wilson, 2010). Por ser um campo recente, mais pesquisas são necessárias e seus métodos e técnicas recém-elaborados devem ser mais bem examinados a fim de legitimar ou não a sua função e utilidade na caixa de ferramentas da ciência.
Entropia genética na especiação
E o que dizer dos custos causados quando realmente ocorre a especiação? Outro grande dificultador para o modelo gradualista é que ele é neutro em relação à melhora ou piora do processo de especiação, embora assuma que a deriva genética cause perdas de variação genética em populações pequenas. O modelo de muitos ancestrais básicos sepultados num catastrofismo recente (SRABUC), por sua vez, munido da proposta da entropia genética, defende que a especiação resulta na perda de informação genética e na consequente degeneração do genoma devido às adaptações como recursos evasivos, perigosos, desgastantes para as populações (Ariza, 2007; Sanford, 2014; Crabtree, 2013a; Crabtree, 2013b). Outras evidências corroboram o modelo SRABVSURCno sentido de sugerir que essa perda de informação genética devido a mutações deletérias em humanos tenha se dado recentemente, entre 5.000 a 10.000 anos atrás (Fu, 2013).
A entropia na informação genética é cada vez mais evidente em tudo que se observa: na deriva genética, na seleção/eliminação, nas mutações, nas complexidades imunológicas desgastantes, entre outros mecanismos. Em humanos, por exemplo, as estimativas atuais é a de que ocorram entre 100-200 novas mutações por indivíduo a cada geração (Nachman e Crowell, 2000; Dolgin, 2009; Lynch, 2010). Destas, os dados variam entre 1-15% de mutações deletérias que causariam a perda direta de informação genética em humanos a cada geração (Nachman e Crowell, 2000; Lynch, 2010; Eyre-Walker e Keightley, 1999;Shabalina et AL., 2001; Keightley, 2012).
Em relação ao fitness, um estudo publicado em 1997 estimou entre 1-2% a taxa de perda da aptidão humana, ou seja, um custo genético alto que faz com que a humanidade se degenere a cada geração devido o esgotamento dos recursos adaptativos utilizados para manter sua variedade genética (Crow, 1997). Em 2010, por sua vez, outro estudo estimou que a aptidão humana está em declínio em 3-5% por geração (Lynch, 2010). Também, outro estudo simulando numericamente o acúmulo de mutações deletérias, demonstra além de a maior parte das mutações estarem a esquerda, (-0.001), ou seja, tendência de serem deletérias, “mutações ruins”), elas também estão acumulando, em uma zona chamada de “Zona de Não-Seleção”, por serem “quase-neutras” essas mutações vem se acumulando ao longo das gerações causando dezenas de “erros ortográficos” no genoma populacional. (Sanford et al 2008)
Figura 2 - Distribuição simulada do acúmulo e frequência de mutações deletérias ao longo de gerações e seus efeitos na aptidão populacional.

Gráfico em vermelho mostra distribuição natural de mutações onde não ocorre seleção. As colunas em verde, mostram as mutações atuais que estão se acumulando, ou seja, dominantes. Outras colunas são mutações menores, recessivas que vem se acumulando à medida que chegam a zona quase neutra, próxima à 0.0.
Sucessivas Derivas Empobrecedoras
O zoólogo holandês Duyvene de Wit descreveu perfeitamente esse processo de empobrecimento genético ao afirmar que:
“quando uma população marginal abre caminho para um novo habitat, ela não pode levar consigo todos os genes da população materna, mas sim apenas parte deles. Cada nova raça ou espécie que se origina de outra possui, portanto, um pool gênico mais pobre. Conseqüentemente, a perda de substância do ‘pool’ gênico é o preço que cada raça ou espécie tem de pagar pelo privilégio de existir. Se o processo de especiação ocorrer repetidas vezes consecutivamente, surgirão por fim, espécies cujos ‘pools’ gênicos estão tão completamente esgotados que bastam alterações insignificantes nas condições ambientais para que elas venham a se extinguir. Esforços para se adaptar a modificações ambientais como resultado de possibilidades insuficientes de recombinação levam, por fim, a um estado genético mínimo. Se este limite mínimo for ultrapassado, não haverá mais possibilidade de sobrevivência. Por essa razão, o destino trágico e irrevogável das espécies ou raças especializadas, extremamente adaptadas, é a morte genética” (Kahle, 1999, p.87; Junker e Scherer, 1996).
Portanto, a proposta SRABVSURC é razoável e se torna cada vez mais corroborada pelos dados científicos ao afirmar que os seres vivos do passado eram mais flexíveis evolutivamente , tinham maior fitness adaptativo, eram saudáveis e sem acúmulo de carga negativa de mutações deletérias e não eram aptos em relação aos problemas de hoje, eram sem os defeitos oriundos das adaptações forçadas e das pressões seletivas distintas a fim de que viessem a subsistir. Vale lembrar que sub-especiações, quanto mais verificáveis em tempo real, mais provam a bio-modificação em curto período de tempo, tal como prevê o presente modelo neocatastrofista de impactos de asteroides.
Registro fóssil e a ausência de diversificação de espécies
Especiação proporcional ao comportamento biodiferenciador observado está ausente pelo fato de existir apenas cerca de 250-300.000 tipos fósseis se repetindo dentre as estimativas de trilhões ou inumeráveis amostras, em todos os estratos geológicos até o pleistosceno (Woodmorappe, 2000; Sadava et al., 2009; Loceye Lennon, 2016). Este fato indica a ausência de sequências durante o processo de especiação quando contrastada com as estimativas de 8,7 milhões de espécies vivas atualmente (Mora et al., 2011). Este salto indica uma catástrofe no meio , assim como temos a obviedade de inicio de camadas sedimentares trazendo rica informação fóssil no ediacara/cambriano e todo o diferencial das camadas do fanerozoico com as pré-cambrianas, tudo nos fala de uma gigantesca catástrofe que iniciou sua deposição ali .
Em outras palavras, a ausência de “diversidade” no registro fóssil revela que as diferenciações ocorrem mais no presente (milhões de espécies) do que em toda coluna geológica (com apenas 3 centenas de milhares). Este episódio indica um tempo em que os seres vivos não precisavam se sub-especiar (bio-modificar) e se adaptar com frequência pra sobreviver, porque viviam em um ambiente mais propício para a vida, estável, assim como a PMS requer.
Evidências recentes apontam que o início da diversificação de alguns gêneros de plantas consideradas “fósseis vivos”, por exemplo, ocorreu ao mesmo tempo ao redor do mundo e em um período muito mais recente do que se supunha, revelando conflitos na proposta gradualista das espécies (Nagalingum et al., 2011). Ademais, foi relatado que essa rápida diversificação teve como causa uma grande mudança climática. Outro exemplo semelhante vem de Ernest Mayr :
“In evolutionary biology we have species like horseshoe crabs. The horseshoe crab goes back in the fossil record over two hundred million years without any major changes. So obviously they have a very invariant genome type, right? Wrong, they don't. Study the genotype of a series of horseshoe crabs and you'll find there's a great deal of genetic variation. How come, in spite of all this genetic variation, they haven't changed at all in over two hundred million years while other members of their ecosystem in which they were living two hundred million years ago are either extinct or have developed into something totally different? Why did the horseshoe crabs not change? That's the kind of question that completely stumps us at the present time.”
Aliás, são vários os motores adaptacionais e modificacionais integrados reconhecidos atualmente. O fato de eles existirem nos seres vivos, nos permite sugerir que as bio-modificações nunca deixaram de existir, nunca e em hipótese alguma não podemos dizer que evolução e flexibilidade nos seres vivos é fato somente hoje.
Outro padrão do registro fóssil parece sugerir mais o movimento de segregação e estratificação espontânea que (Dilly et al, 2015: Berthaut, 1986, 1988, 2002, 2004, 2010, 2011, 2012, 2013, 2014; Lalomov, 2007, 2013; Julien, P.Y, 1993), reproduzem em laboratório . Um deles é o desaparecimento coordenados: “Desaparecimentos coordenados . Um grande número de espécies fósseis podem desaparecer do registro geológico em um nível estratigráfico específico (ver Figura 3). O desaparecimento nunca está completo, mas existem vários exemplos em que as estimativas indicam que mais de 50% das espécies desaparecem no mesmo nível estratigráfico. 6Os limites entre níveis estratigráficos são frequentemente identificados com base em desaparecimentos coordenados. O maior exemplo disso é o desaparecimento de quase metade das famílias (ver Figura 3) e cerca de 95% de todas as espécies no topo do Paleozoico. Dinossauros e muitos outros grupos de répteis e invertebrados marinhos desaparecem do registro no topo do Mesozóico. Outros exemplos de desaparecimentos coordenados em larga escala ocorrem na parte superior do ordoviciano, perto do topo do Devoniano e do topo do Triássico. O desaparecimento coordenado é um padrão de subtração. Não foram relatadas tendências sustentadas neste padrão”.(Gibson, 1996)
Figura 3 - Padrão estratigráfico do número de famílias de invertebrados marinhos representados pelos fósseis em cada nível estratigráfico. (Após Sepkoski 1993, ver Nota 6.)
Nível Estratigráfico: V = Vendian; Cm = Cambriano; O = ordovicêutico; S = Siluriano; D = Devoniano; C = Carbonífero; P = Permiano; Tr = Triássico; J = Jurássico; K = Cretáceo; T = Terciário. Pontos marcados como 1-5 representam as cinco maiores "extinções em massa"
A segregação estratigráfica espontânea de corpos e sedimentos comuns, explica uma certa alternância entre presença de muitos fósseis de uma categoria seguido de falta dela (o que é interpretado como extinção em massa) .
O PMS gera muita necessidade de deduções e modelos modelos matemáticos, simulações macroevolutivas em computadores, entre inumeráveis justificativas, que como esforços , como se referiu o Dr Tamborini em relação a paleontologia idiográfica, "são científicos em um sentido puramente descritivo, mas são completamente inúteis para investigações biológicas. Finalmente” (Tamborini, 2015).
Alegações que em 90% dos 544 milhões de anos totais do Fanerozóico, as bio-modificações se estagnaram, repetindo na coluna geológica as mesmas espécies, e só recentemente nas camadas mais atuais (superiores) é que houve biodiversificação em larga escala (radiação) (Gould, 1981) dão um exemplo de descrições científicas terem maior peso do que os dados em si. Além destas justificativas em relação a PMS, temos gigantesca lista de ad hocs justificadores de claros anacronismos geocronológicos como por exemplo, tecidos moles e suposta inesperada preservação de proteínas de animais fossilizados, datados entre 60-120 Ma; onde os autores em vez de questionarem as intocáveis datações absolutas, resolvem buscar formas de justificar incriveis preservações de frágeis proteínas, outro exemplo vem da propria radiometria datacional , com a presença inesperada de carbono 14 datável em diamantes “incontamináveis” de 300 Ma (Baumgardner, 2005);, e quase uma centena de perspectivas e métodos datacionais , que são esquecidos, pela preferência daquilo que combina com o tempo do paradigma geocronológico vigente. Ver TABELA 2.
Figura 4 - Fluxo das interpretações geocronológicas quanto ao PMS, tecidos moles, carbono 14 , onde a geocronologia assumida obriga pesquisadores a fabricarem justificativas ad hoc a direita do que interpretarem o fato em si .

Calcula-se que C14 numa amostra tenha desaparecido totalmente após 100-250 mil anos , já que sua meia vida é de apenas 5730 anos , fato que leva muitos ao dogma de que não se testa coisas supostamente de Ma com C14. No entanto, desafiando esta convenção, foram feitos milhares de testes da equipe do geofísico Dr John Baumgardner no çlaboratorio de Los Álamos, Texas, os quais demonstraram que rochas de 300-500 milhões de anos continham quantificações de teor datável de C14; diante de ad hocs justificando tal anomalia , os críticos recorreram a contaminação, então eles testaram até mesmo em diamantes “incontamináveis” ( a possibilidade de contaminação em diamantes chega a ser desprezível) dando semelhantes resultados e confrontando a geocronologia convencional. (Baumgardner, 2005)
Dr. Tom Kemp, curador das coleções zoológicas do Museu de História Natural da Universidade de Oxford, por exemplo, fez a seguinte admissão: “como é agora bem conhecido, a maioria das espécies fósseis aparecem instantaneamente no registro fóssil, persistem por alguns milhões de anos praticamente inalterados, e apenas desaparecem abruptamente” (Kemp, 1985, p. 67).
Darwin sugeriu que os seres vivos surgiam por evolução gradativa, e esperava que um dia o registro fóssil confirmasse sua predição, porém não foi o que ocorreu: Darwin estava errado. Dia após dia, os inúmeros fósseis escavados por todo o mundo têm refutado a hipótese de mudança gradual em camadas sobrepostas supostamente cronológicas. Ao contrário do que Darwin esperava, os dados recentes revelam padrões de aparecimento súbito (explosões) seguido por longos períodos de pouca mudança.
O famoso paleontólogo evolucionista Niles Eldredge admitiu na revista New Scientist que:
"os paleontólogos desde Darwin têm feito buscas (em grande parte em vão) de sequências de séries graduadas de fósseis que se destacam como exemplos do tipo de transformação global de espécies que Darwin imaginou como o produto natural do processo evolutivo. Poucos viram qualquer razão para duvidar − embora seja um fato surpreendente que [...] a maioria das espécies permanecem reconhecidamente as mesmas, praticamente inalteradas ao longo de sua ocorrência em estratos geológicos de várias idades" (Eldredge, 1986, p.55).
A ausência de fósseis intermediários é demasiado óbvia para os evolucionistas encobrirem por mais tempo. O biólogo evolutivo Dr. David Woodruff da Universidade da Califórnia expressou na revista Science o desapontamento de evolucionistas relativo à ausência no registro fóssil de formas transicionais: "as espécies fósseis permanecem inalteradas durante a maior parte de sua história e o registro não consegue conter um único exemplo de uma transição significativa." (Woodruff, 1980, p.716).
Portanto, diante de tantas admissões, finalmente concordamos com a bióloga evolucionista Lynn Margulis quando diz que:
“não há nenhum gradualismo no registro fóssil. [...] O ‘equilíbrio pontuado’ foi inventado para descrever a descontinuidade no surgimento de novas espécies. [...] Os críticos, [incluindo os críticos SRABUCS], estão certos nas críticas que fazem. [...] Os biólogos evolucionistas acreditam que o padrão evolucionário é uma árvore. Não é!” (Teresi, 2011).
Exemplos de especiação rápida
Existem diversos relatos de surgimento de novas “espécies” em períodos que variam de dezenas há milhares de anos. Segundo essas informações, a especiação é um fenômeno que não necessita de milhões de anos para acontecer. O biólogo norte-americano Dr. James Gibson, Diretor do Instituto de Pesquisa em Geociência (GRI, na sigla em inglês), instituição afiliada à Universidade de Andrews (EUA), mantida pela IASD, descreveu um exemplo de especiação em tempo real: “uma nova espécie de copépode [crustáceos] formou-se no Mar Salton no sul da Califórnia em menos de 30 anos.” (Johnson, 1953; Gibson, 2002).
Estudos diversos têm relatado que apenas entre 10 a 36 anos as populações diferentes de lagartos sofreram alterações morfológicas significativas o suficiente para serem consideradas novas “espécies” (Morrell, 1997;Herrel et al., 2008). Outros exemplos conhecidos de especiação em tempo real incluem bactérias (Shikano et al., 1990), moscas (Huey et al., 2000), tentilhões (Grant e Grant, 2006), rãs (Hoskin et al., 2005), escaravelhos (Halliburton e Gall, 1981) e plantas (Groves e Groves, 1880; Foucaud, 1897; Marchant, 1963).
Dr. Gibson forneceu ainda exemplos de especiação em tempo histórico-arqueológico: “uma população de macacos verdes viveu na ilha de St Kitts no Caribe por menos de 100 anos, mas desenvolveu aspectos morfológicos equivalentes a uma nova espécie.” (Ashton et al., 1979; Gibson, 2002). “O Havaí não tinha bananeiras até cerca de 1000 anos atrás, no entanto há mariposas havaianas nativas que só se alimentam de bananeiras. Estas novas espécies surgiram em menos de 1000 anos.” (Zimmerman, 1960; Gibson, 2002). Diante desses exemplos, Dr. Gibson conclui: “a capacidade de mudanças rápidas está confirmada tanto por experimentação como por observação da natureza” (Gibson, 2002).
TESTEMUNHO ARQUEOLÓGICO
Mais de uma centena de relatos arqueológicos apresentam cenas apavorantes do que ocorreu recentemente na terra . A maioria destes relatos não apresentam aspectos simbólicos, míticos, mas demonstram ser uma descrição literal de um fato dando detalhes as vezes técnicos e matemáticos. Um relato de um livro sagrado dos Maias , o "Chilam Balaam", parece descrever cenas do que poderíamos pensar em termos de consequências de impactos de asteroides na terra, da seguinte forma:
"Ah Mucencab veio e obscureceu a face dos Céus... a Terra começou a despertar. Ninguém sabia o que ia acontecer. De repente, fogos subterrâneos irromperam subindo Firmamento adentro e choveu fogo do alto, e despencaram cinzas, e pedras e árvores foram lançadas para baixo, e madeiras e pedras se despedaçaram umas contra as outras. Então o Céu foi agarrado e afastado à força. A face do Céu foi fustigada de um lado para o outro e atirada para trás... as pessoas foram todas esmigalhadas; seus corações pararam enquanto elas ainda viviam. Então elas foram enterradas nas areias, no mar. Numa enorme e repentina torrente de água, a Grande Serpente foi arrebatada do Céu. O Firmamento caiu e a Terra afundou quando os quatro deuses, os Bacabs, apresentaram quem trouxe a destruição do mundo." . Os Incas disseram : Incas "Um dia, um grupo de estrangeiros chegou para avisar que os orgulhosos habitantes daquela cidade seriam destruídos pelo terremoto, pela inundação e pelo fogo. A maioria deles riu dos estrangeiros. Contudo, alguns sacerdotes da cidade ouviram o aviso e foram viver em lugares afastados no topo das montanhas. Algum tempo depois, uma nuvem vermelha apareceu no horizonte.Ela cresceu e cobriu toda a área, e sua vermelhidão era tão forte que iluminava o céu noturno. De repente, com um brilho e um estrondo, um terremoto destruiu muitos edifícios da cidade, e uma chuva vermelha começou a cair.Outros terremotos e mais chuva vieram em sequência, e uma inundação logo cobriu toda a cidade arruinada" Outro relato dos maias esse retirado do Popul Vuh: "Então as águas foram agitadas pela vontade de Hurakán, e uma grande inundação desabou sobre as cabeças dessas criaturas... Elas foram tragadas e uma nebulosidade resinosa desceu do céu... a face da Terra ficou sombria e uma pesada chuva que tudo escurecia começou a cair — chuva de dia e chuva de noite... Ouviu-se um grande barulho acima de suas cabeças, como se produzido pelo fogo. Então se viram homens correndo, empurrando-se, cheios de desespero; quiseram subir em suas casas, e as casas, desmoronando, caíram por terra; quiseram trepar nas árvores, e as árvores fizeram chacoalharam à sua frente"
Conclusão
Padrões morfológicos em torno do táxon gênero (PMTG) se identificam com fósseis ancestrais básicos sepultados num catastrofismo recente (SRABUC), pois este modelo respeita o fato da especiação rápida observada, que exigiria uma variabilidade proporcional muito grande no registro fóssil, caso representasse a evolução em milhões de anos (PMS), e isto além de não existir tal variação taxonômica, ainda se confirma com estase morfológica, repetição das mesmas espécies e 4229 gêneros de fósseis vivos, quadro este que reflete um sepultamento repentino dos seres vivos do planeta e não uma suposta história evolutiva da vida de Ma. Relatos de especiação na história e arqueologia, capazes de justificar toda biodiversidade em pouco tempo, nos mostram que deveríamos ter muito mais variabilidade no registro fóssil do que há, o que, dado o fato da evolução ser verificada, podemos certificar que não houve tempo para que ela atuasse. A explosão cambriana, a ausência de variabilidade amostral fóssil (estase morfológica fóssil e pobreza taxonômica) e falta de radiação especiativa no registro fóssil (diversificação) até o pleistoceno, nos conta uma história de um período de: 1) surgimento rápido e pronto de formas de vida , 2) permanência morfológica com alto número de espécies em ambiente estável (repetição fóssil sem pressões ambientais evolutivas e adaptativas, e sem grandes catástrofes intercalando tempo), 3) desastre soterrador de população viva evidenciada pelas repetições de mesmas espécies fósseis (o que descaracteriza a pontualidade), 4) presença de diversas espécies diferentes unidas no registro fóssil, fósseis de imensos vertebrados completos continentais(o que caracteriza desastre de grande magnitude e altas taxas de sedimentação), e 5) mudança drástica no ambiente gerando a radiação das espécies (diversificação) nas camadas recentes de forma amostral (fósseis recentes) e nas milhões de espécies na biodiversidade atual. “Enquanto há 200 anos, os naturalistas achavam que havia talvez cinco ou dez mil espécies na Terra, estimativas atuais... coloque o valor em dez ou 20 milhões. Costuma-se supor que a vida nunca tenha sido mais diversificada do que é hoje”. (Benton et al, 2007) Estes pontos levantados, questionam a geocronologia , e não buscam , como muitos de artigos científicos o fazem, justificar com ad hocs as anomalias anacrônicas do que se esperaria encontrar nos fatos paleontológicos, mas destacam inteerpretações mais independentes da geocronologia convencional, sugerindo assim amplo questionamento das datações de alta escala de tempo , chamadas de “absolutas” e em especial, da hipótese uniformitarianista de que nada acelerou o decaimento radioativo envelhecendo rochas durante os supostos bilhões de anos aferidos, que ainda imperam por décadas sobre o pensamento dinâmico e científico moderno, mesmo possuindo talvez centenas de métodos e aperfeiçoamentos para acelerar decaimento radioativo e digamos assim, envelhecer as rochas com voltagens e ambientes que representam uma irrisória comparação com o que ocorreu quando bólidos imensos cairam sobre a terra, onde aproximadamente 99,8% foram apagados pela dinâmica tectônica, marítma , erosiva e sedimentar da terra.
A estase morfológica no registro fóssil só pode representar o sepultamento de uma população em um só momento , qualquer interpretação que avançar um milímetro sequer desta realidade, contrariará tudo que conhecemos sobre evolução e plasticidade adaptativa das espécies ao longo do tempo e a hipótese de uma chuva de asteroides parece ser suficiente para poder não somente explicar o PMS, mas quase todas as anomalias geoquímicas e geocronológicas da terra. Milhares de estudos poderão ser desenvolvidos a partir destas poucas reflexões introdutórias que fizemos aqui.
REFERÊNCIAS
Goodings, CR & Brookfield, ME, 1992.Movimentos transcorrentes proterozóicos ao longo do linfonodo Kapuskasing (Província Superior, Canadá) e sua relação com as estruturas circundantes. Earth-Science Reviews, 32: 147-185.
Bleeker, W. e Pilkington, M., 2004.O Arco Nastapoka de 450 km de diâmetro: a cicatriz de impacto preservada mais antiga e maior da Terra? Programa com resumos - Associação Geológica do Canadá; Associação Mineralógica do Canadá: Reunião Anual Conjunta, 2004, Vol. 29, pp. 344.
Beals, CS, 1968. Sobre a possibilidade de uma origem catastrófica para o grande arco do leste da baía de Hudson. Em: Beals, CS (Ed.), Pp. 985-999. Ciência, História e Hudson Bay , Vol. 2, Ministério das Minas e Recursos da Energia, Ottawa.
1. Genesis 1:1; 7:11,12
2. Chilam Balam : (Chumayel). Barcelona: Linkgua Ediciones. 2008. ISBN 9788498971439. OCLC 502270722
3. Aaron M (2014). Baraminological Analysis of the Caseidae (Synapsida: Pelycosauria). Journal of Creation Theology and Science Series B: Life Sciences. 4:19-22.
4. Albrecht C, Wilke T (2008). Ancient Lake Ohrid: biodiversity and evolution. Hydrobiologia. 615(1):103-140.
5. Alisson E (2013). Arqueologia ajudará a desvendar origem da biodiversidade amazônica. Agência FAPESP. Disponível em: http://agencia.fapesp.br/arqueologia_ajudara_a_desvendar_origem_da_biodiversidade_amazonica/16937/
6. Ariza LM (2007). Evolution in a Petri Dish. Scientific American. 297(6):36. Disponível em: http://www.scientificamerican.com/article/evolution-in-a-petri-dish/
7. Ashton EH, Flinn RM, Griffiths RK, Moore WJ (1979). The results of geographic isolation on the teeth and skull of the Green monkey (Cercopithecus aethiopssabaeus) in St. Kitts – a multivariate retrospect. Journal of Zoology. 188(4):533-555.
8. Aw Kinderewitsch e Li Kitscha: Polewaja suschtschnost jadernoi fisiki - campo de Física Nuclear, Kiev: EKMO, 2003, páginas 263 -30
Basener, William F.; Sanford, John C. (7 de novembro de 2017). «The fundamental theorem of natural selection with mutations». Journal of Mathematical Biology(em inglês): 1–34. ISSN 0303-6812. doi:10.1007/s00285-017-1190-x
Barton, PJ, Grieve, RAF, Morgan, JV, Surendra, AT, Vermeesch, PM, Christeson, GL, Gulick, SPS e Warner, MR, Imagens sísmicas da folha de derretimento de impacto Chicxulub e comparação com a estrutura de Sudbury; em: Gibson, RL e Reimold, WU (Eds.), grandes impactos de meteoritos e evolução planetária IV , papel especial GSA 465, pp. 103-113, 2010.
9. Baumgardner, Jonh (2005). «Radioisotopes and the Age of the Earth, Volume II». Institute for Creation Research. Consultado em 10 de setembro de 2017
10. Behe, M. June 1997). «Darwinism and design». Trends in Ecology & Evolution. 12 (6). 229 páginas. ISSN 0169-5347. PMID 2. (J1238050
11. Behe, Michael J. (February 2009). «Waiting longer for two mutations». Genetics. 181 (2): 819–820; author reply 821–822. ISSN 0016-6731. PMID 19189948. doi:10.1534/genetics.108.098905
12. Bell G (2013). Evolutionary rescue and the limits of adaptation. Philos Trans R Soc Lond B Biol Sci. 368(1610):20120080.
13. Benedict, John C.; Smith, Selena Y.; Specht, Chelsea D.; Collinson, Margaret E.; Leong-Škorničková, Jana; Parkinson, Dilworth Y.; Marone, Federica (2016). «Species diversity driven by morphological and ecological disparity: a case study of comparative seed morphology and anatomy across a large monocot order». AoB PLANTS. 8. ISSN 2041-2851. PMID 27594701
14. Benton, Michael J.; Emerson, Brent C. (1 de janeiro de 2007). «How Did Life Become so Diverse? the Dynamics of Diversification According to the Fossil Record and Molecular Phylogenetics». Palaeontology (em inglês). 50 (1): 23–40. ISSN 1475-4983. doi:10.1111/j.1475-4983.2006.00612.x
15. Berthault, G. : “Orogenesis: Cause of sedimentary formations” – Kazan Golovkinsy Stratigraphic Meeting, 2014, pp.19-20
16. Berthault, G. : “Orogenesis: cause of sedimentary formations” – “Open Journal of Geology“ ISSN 2161-7570.Vol 3, Number 28, April 2013.
17. Berthault G. : “Towards a Refoundation of Historical Geology” – “Georesources” 1(12) 2012, p.38, 39
18. Berthault, G., Lalomov, A. V. and Tugarova, M. A. : “Reconstruction of paleolithodynamic formation conditions of Cambrian-Ordovician sandstones in the Northwestern Russian platform” – “Lithology and Mineral Resources, 2011, Volume 46, Number 1, 60-70” (Springer Publishing site)
19. Berthault, G., Veksler A.B., Donenberg V.M. , Lalomov A. : “RESEARCH on EROSION OF CONSOLIDATED and semi-consolidated SOILS BY HIGH SPEED WATER FLOW” Izvestia.VNIIG., 2010, Vol. 257, pp.10-22. – (Russian original.)
20. Berthault, G : “Sedimentological Interpretation of the Tonto Group Stratigraphy (Grand Canyon Colorado River)” , Lithology and Mineral Resources 2004, Vol. 39, No 5. October 2004.
21. Berthault G., “Analysis of Main Principles of Stratigraphy on the Basis of Experimental Data”, Litol.Polezn.Iskop.2002, vol 37, no.5,pp 509-515 (Lithology and Mineral resources 2002 (fac-similé) (Engl.Transl.), vol.37, no.5, pp442-446), Journal of the Academy of Sciences of Russia.
22. Berthault, G., “Sedimentation of a Heterogranular Mixture. Experimental Lamination in Still and Running Water”, Compte rendu de l’Académie des Sciences 1988, vol. 306, Serie II, pp. 717–724.
23. Berthault, G., “Sedimentologie: Expériences sur la lamination des sédiments par granoclassement périodique postérieur au dépôt. Contribution a l’explication de la lamination dans nombre de sédiments et de roches sédimentaires”., Compte rendu de l’Académie des Sciences de Paris 1986 , vol. 303, Ser., 2, no. 17, pp. 1569-1574.
24. Bhullar BS (2017). Evolution: Catastrophe triggers diversification. Nature. 542 (7641):304-305.
25. Bosch, Fritz (1999). «Setting a Cosmic Clock with Highly Charged Ions». Physica Scripta (em inglês). T80 (A). ISSN 1402-4896. doi:10.1238/physica.topical.080a00028
26. Brand LR, Tang T (1991). Fossil vertebrate footprints in the Coconino Sandstone (Permian) of northern Arizona: Evidence for underwater origin. Geology. 19(12):1201-1204.
27. Brawn, Dan (2013). «The Origin of Earth's Radioactivity». Consultado em 24 de setembro de 2017
28. Bunch, Ted E.; Hermes, Robert E.; Moore, Andrew M. T.; Kennett, Douglas J.; Weaver, James C.; Wittke, James H.; DeCarli, Paul S.; Bischoff, James L.; Hillman, Gordon C. (10 de julho de 2012). «Very high-temperature impact melt products as evidence for cosmic airbursts and impacts 12,900 years ago». Proceedings of the National Academy of Sciences of the United States of America. 109 (28): E1903–1912. ISSN 1091-6490. PMID 22711809. doi:10.1073/pnas.1204453109
29. Carroll, R. L. (1992). «The Primary Radiation of Terrestrial Vertebrates». Annual Review of Earth and Planetary Sciences. 20 (1): 45–84. doi:10.1146/annurev.ea.20.050192.000401
30. Catalogue of Life (2016). Species 2000 & ITIS Catalogue of Life, 2016 Annual Checklist. Disponível em: http://www.catalogueoflife.org/col/info/ac
31. Cavanaugh DP, Sternberg RV (2004). Analysis of morphological groupings using ANOPA, a pattern recognition and multivariate statistical method: A case study involving centrarchid fishes. Journal of Biological Systems. 12(2):137–67.
32. Cavanaugh DP, Wood TC (2002). A Baraminological Analysis of the Tribe Heliantheaesensulato (Asteraceae) Using Analysis of Pattern (ANOPA). Occas. Papers of the BSG. 17(1):1-11.
33. Chadwick A, Spencer L (2006). Turner Preliminary depositional model for an Upper Cretaceous Edmontosaurusbonebed. Journal of Vertebrate paleontology. 26:49A.
34. Crabtree GR (2013a). Our fragile intellect. Part I. Trends in Genetics. 29(1):1-3.
35. Crabtree GR (2013b). Our fragile intellect. Part II. Trends in Genetics. 29(1):3-5.
36. Crow JF (1997). The high spontaneous mutation rate: is it a health risk? Proc Natl AcadSci U S A. 94(16):8380-6.
37. Darwin C (1861). On the Origin of Species. 3. Ed. London: Murray. capítulo 14.
38. de Paula MO (2009). Podem os criacionistas aceitar a origem de novas espécies? Earth History Research Center. Disponível em: http://origins.swau.edu/papers/evol/marcia1/defaultp.html
39. de Queiroz K (2005). Ernst Mayr and the modern concept of species. Proc Natl AcadSci U S A. 102 Suppl 1:6600-7.
40. Dilly, R., Berthault, G.: “Orogenesis: Cause of sedimentary formations” – The Russian Academy of Sciences scientific Council on Lithology and Minerals in Sedimentary Formations – VIII All-Russian Lithological Meeting (Moscow, 27-30 October 2015), Tome II, pp. 162-164
41. Dolgin E (2009). Human mutation rate revealed. Nature News (27 Ago. 2009). Disponível em:http://www.nature.com/news/2009/090827/full/news.2009.864.html
42. Donnelley, Strachan (July 2002). «Natural responsibilities. Philosophy, biology, and ethics in Ernst Mayr and Hans Jonas». The Hastings Center Report. 32(4): 36–43. ISSN 0093-0334. PMID 12362522
43. Eakin CM (2014). Lamarck was partially right—and that is good for corals. Science. 344(6186):798-9.
44. Eldredge N, Stanley SM (1984). Living Fossils: Introduction to the Casebook. New York/Berlin: Springer-Verlag pp. 266-270.
45. Eldredge N (1986). Progress in Evolution? New Scientist. 110:54-57.
46. Eyre-Walker A, Keightley PD (1999). High genomic deleterious mutation rates in hominids. Nature. 397(6717):344-7.
47. Ezard, Thomas H. G.; Quental, Tiago B.; Benton, Michael J. (5 de abril de 2016). «The challenges to inferring the regulators of biodiversity in deep time». Phil. Trans. R. Soc. B(em inglês). 371 (1691). 20150216 páginas. ISSN 0962-8436. PMID 26977058. doi:10.1098/rstb.2015.0216
48. Grant, B. Rosemary; Grant, Peter R. (3 de março de 2017). «Watching speciation in action». Science (New York, N.Y.). 355 (6328): 910–911. ISSN 1095-9203. PMID 28254901. doi:10.1126/science.aam6411.
49. Folha de S. Paulo (2006). Pássaros exibem seleção natural em tempo real. Folha de São Paulo (14/07/2006). Disponível em: http://www1.folha.uol.com.br/folha/ciencia/ult306u14848.shtml
50. Fondon JW, Garner HR (2004). Molecular origins of rapid and continuous morphological evolution. Proc Natl AcadSci U S A. 101(52):18058-63.
51. Foote, Mike (1993/04). «Discordance and concordance between morphological and taxonomic diversity». Paleobiology. 19 (2): 185–204. ISSN 0094-8373. doi:10.1017/S0094837300015864
52. Foucaud (1897). Un Spartinainédit. Ann SocSci Nat Char Inf. 32:220–222.
53. Frair W (2000). Baraminology: Classification of Created Organisms. CRSQ Quarterly. 37(2):82-91.
54. Freeman S, Herron JC (2009). Análise evolutiva. 4. ed. orto Alegre: Artmed. 848p.
55. Fu W, et al (2013). Analysis of 6,515 exomes reveals the recent origin of most human protein-coding variants. Nature. 493(7431):216-20.
56. Futuyma, Douglas J. (July 2010). «Evolutionary constraint and ecological consequences». Evolution; International Journal of Organic Evolution. 64(7): 1865–1884. ISSN 1558-5646. PMID 20659157. doi:10.1111/j.1558-5646.2010.00960.x
57. Furness, Andrew I.; Lee, Kevin; Reznick, David N. (June 2015). «Adaptation in a variable environment: Phenotypic plasticity and bet-hedging during egg diapause and hatching in an annual killifish». Evolution; International Journal of Organic Evolution. 69 (6): 1461–1475. ISSN 1558-5646. PMID 25908306. doi:10.1111/evo.12669
58. Gentry, R.V. 1970. "Giant Radioactive Halos: Indicators of Unknown Alpha-Radioactivity?" Science 169, 670.
59. Gentry, R.V. 1971. "Radiohalos: Some Unique Pb Isotope Ratios and Unknown Alpha Radioactivity." Science 173, 727.
60. Gentry, R.V. 1973. "Radioactive Halos." Annual Review of Nuclear Science 23, 347. Acessado em 24/09/2017 em http://www.robertvgentry.com/
61. Gentry, R.V. 1974. "Radiohalos in Radiochronological and Cosmological Perspective." Science 184, 62. Acessado em 24/09/2017 em http://www.robertvgentry.com/
62. Gentry, R.V. 1977. "Mystery of the Radiohalos." Research Communications NETWORK, Breakthrough Report, February 10, 1977. Acessado em 24/09/2017 em http://www.robertvgentry.com/
63. Gentry, R.V. 1978a. "Are Any Unusual Radiohalos Evidence for SHE?" International Symposium on Superheavy Elements, Lubbock, Texas. New York: Pergamon Press.
64. Gentry, R.V. 1978b. "Implications on Unknown Radioactivity of Giant and Dwarf Haloes in Scandinavian Rocks." Nature 274, 457.
65. Gentry, R.V. 1978c. "Reinvestigation of the α Activity of Conway Granite." Nature 273, 217. PDF
66. Gentry, R.V. 1979. "Time: Measured Responses." EOS Transactions of the American Geophysical Union 60, 474. Acessado em 24/09/2017 em http://www.robertvgentry.com/
67. Gentry, R.V. 1980. "Polonium Halos." EOS Transactions of the American Geophysical Union 61, 514.
68. Gentry, R.V. et al. 1973. "Ion Microprobe Confirmation of Pb Isotope Ratios and Search for Isomer Precursors in Polonium Radiohalos." Nature 244, 282.
69. Gentry, R.V. et al. 1974. "'Spectacle' Array of Po-210 Halo Radiocentres in Biotite: A Nuclear Geophysical Enigma." Acessado em 24/09/2017 em http://www.robertvgentry.com/
70. Gentry, R.V. et al. 1976a. "Radiohalos and Coalified Wood: New Evidence Relating to the Time of Uranium Introduction and Coalification." Science 194, 315 . Acessado em 24/09/2017 em http://www.robertvgentry.com/
71. Ghalambor, Cameron K.; Hoke, Kim L.; Ruell, Emily W.; Fischer, Eva K.; Reznick, David N.; Hughes, Kimberly A. (17 de setembro de 2015). «Non-adaptive plasticity potentiates rapid adaptive evolution of gene expression in nature». Nature (em inglês). 525 (7569): 372–375. doi:10.1038/nature15256
72. Gibson LJ (2002). A similaridade biológica e a teoria da evolução. In: 4º Encontro Nacional de Criacionistas, 24 a 28 de janeiro de 2002, São Paulo. Anais... São Paulo: Centro Universitário Adventista, campus São Paulo.
73. Gibson, L. J. --- Fossil Patterns: A Classification and Evaluation». www.grisda.org. Consultado em 10 de setembro de 2017
74. Gorodezki V, 2005. Deactivation unit for radioactive materials, comprises capacitor with curved mantle surfaces, connected to controllable generator DE 10354251 A1.Acessado em 24/09/2017 em http://www.google.com.na/patents/DE10354251A1?cl=en&hl=pt-BR
75. Gould SJ (1981). Evolution as Fact and Theory, Discover magazine 1981; 2(5):34-37; Reimpresso com permissãoem: Gould SJ. Hen's Teeth and Horse's Toes. New York: W. W. Norton & Company, 1994, pp. 253-262.
76. Grant PR, Grant BR (2006). Evolution of Character Displacement in Darwin's Finches. Science. 313(5784):224-6.
77. Groves H, Groves J (1880). Spartinatownsendiinobis. Rep Bot SocExch Club Br Id. 1:37.
78. Donald R. Lowe, Gary R. Byerly, Frank T. Kyte, Alexander Shukolyukov, Frank Asaro e Alexandra Krull. Astrobiologia. Julho de 2004, 3 (1): 7-48. https://doi.org/10.1089/153110703321632408
79. Halliburton R, Gall GAE (1981). Disruptive selection and assortative mating in Triboliumcastaneum. Evolution. 35:829-843.
80. Herrel A, et al (2008). Rapid large-scale evolutionary divergence in morphology and performance associated with exploitation of a different dietary resource. Proc Natl AcadSci U S A. 105(12):4792-5.
81. Hesslow, L.; Embréus, O.; Stahl, A.; DuBois, T. C.; Papp, G.; Newton, S. L.; Fülöp, T. (23 de junho de 2017). «Effect of Partially Screened Nuclei on Fast-Electron Dynamics». Physical Review Letters. 118 (25). 255001 páginas. ISSN 1079-7114. PMID 28696736. doi:10.1103/PhysRevLett.118.255001
82. Horner JR, Goodwin MB (2009). Extreme Cranial Ontogeny in the Upper Cretaceous Dinosaur Pachycephalosaurus. PLoS One. 4(10):e7626.
83. Hoskin CJ1, Higgie M, McDonald KR, Moritz C (2005). Reinforcement drives rapid allopatric speciation. Nature. 437(7063):1353-6.
84. Huey RB, et al (2000). Rapid Evolution of a Geographic Cline in Size in an Introduced Fly. Science. 287(5451):308-309.
85. I Have A List"». Eye on the ICR (em inglês). 31 de julho de 2011
86. Izumi, Y.; Tomita, H.; Nakayama, Y.; Hayashi, S.; Morishima, K.; Isobe, M.; Cheon, M. S.; Ogawa, K.; Nishitani, T. (November 2016). «Development of fast neutron pinhole camera using nuclear emulsion for neutron emission profile measurement in KSTAR». The Review of Scientific Instruments. 87 (11): 11D840. ISSN 1089-7623. PMID 27910454. doi:10.1063/1.4963866
87. Jerlström P (2000). Is the evolutionary tree turning into a creationist orchard? Journal of Creation. 14(2):11–13.
88. Johnson MW (1953). The copepod Cyclops dimorphus Kiefer from the Salton Sea. American Midland Naturalist. 49:188-192.
89. Julien, P.Y., Lan, Y., and Berthault, G., “Experiments on Stratification of Heterogeneous Sand Mixtures”, Bulletin Société Géologique de France, 1993, vol. 164, no. 5, pp. 649–660.
90. Junker R, Scherer S (1996). Evollução: um livro texto crítico. 4. Ed. Brasília, DF: Sociedade Criacionista Brasileira.
91. Kahle H (1999). Evolution: IrrwegmodernerNaturwissenschaft? 4. Ed. Bielefeld: Mindt, Edeltraud. 200p.
92. Keightley PD (2012). Rates and Fitness Consequences of New Mutations in Humans. Genetics. 190(2):295-304.
93. Kemp TS (1985). A Fresh Look at the Fossil Record. New Scientist. 108(1485):66-67.
94. Kennett, James P.; Kennett, Douglas J.; Culleton, Brendan J.; Aura Tortosa, J. Emili; Bischoff, James L.; Bunch, Ted E.; Daniel, I. Randolph; Erlandson, Jon M.; Ferraro, David (11 de agosto de 2015). «Bayesian chronological analyses consistent with synchronous age of 12,835-12,735 Cal B.P. for Younger Dryas boundary on four continents». Proceedings of the National Academy of Sciences of the United States of America. 112 (32): E4344–4353. ISSN 1091-6490. PMID 26216981. doi:10.1073/pnas.1507146112
95. Kennett, James P.; Kennett, Douglas J.; Culleton, Brendan J.; Aura Tortosa, J. Emili; Bischoff, James L.; Bunch, Ted E.; Daniel, I. Randolph; Erlandson, Jon M.; Ferraro, David (11 de agosto de 2015). «Bayesian chronological analyses consistent with synchronous age of 12,835-12,735 Cal B.P. for Younger Dryas boundary on four continents». Proceedings of the National Academy of Sciences of the United States of America. 112 (32): E4344–4353. ISSN 1091-6490. PMID 26216981. doi:10.1073/pnas.1507146112
96. Kuhn, Joseph A. (2012-1). «Dissecting Darwinism». Proceedings (Baylor University. Medical Center). 25 (1): 41–47. ISSN 0899-8280. PMID 22275784
97. Lacy, Robert C. (1 de agosto de 1987). «Loss of Genetic Diversity from Managed Populations: Interacting Effects of Drift, Mutation, Immigration, Selection, and Population Subdivision». Conservation Biology (em inglês). 1 (2): 143–158. ISSN 1523-1739. doi:10.1111/j.1523-1739.1987.tb00023.x
98. Lalomov, A., Berthault G., Tugarova, M., Isotov V., Sitdikova L.: “Reconstruction of sedimentary conditions of Middle Permian Kama-Ural basin studied by N.A.Golovkinsky” – Kazan Golovkinsy Stratigraphic Meeting, 2014, pp.53-54
99. Lalomov, A. : “Reconstruction of Paleohydrodynamic Conditions during the Formation of Upper Jurassic Conglomerates of the Crimean Peninsula”, Lithology and Mineral Resources, 2007, Vol. 42, No. 3, pp. 268–280
100. Lavoué, Sébastien; Miya, Masaki; Arnegard, Matthew E.; McIntyre, Peter B.; Mamonekene, Victor; Nishida, Mutsumi (7 de abril de 2011). «Remarkable morphological stasis in an extant vertebrate despite tens of millions of years of divergence». Proceedings of the Royal Society B: Biological Sciences. 278 (1708): 1003–1008. ISSN 0962-8452. PMID 20880884.
101. Lee, Kanani K. M.; Steinle-Neumann, Gerd (30 de março de 2008). «Ab-initio study of the effects of pressure and chemistry on the electron-capture radioactive decay constants of 7Be, 22Na and 40K». Earth and Planetary Science Letters. 267 (3): 628–636. doi:10.1016/j.epsl.2007.12.014
102. Levinton JS, Chris MS (1980). A Critique of the Punctuated Equilibria Model and Implications for the Detection of Speciation in the Fossil Record. Syst Zool. 29(2):130–142.
103. Lightner JK (2016). Variation after the Flood. Creation Matters. 21(2):6-7.
104. Locey KJ, Lennon JT (2016). Scaling laws predict global microbial diversity. Proc Natl AcadSci U S A. 113(21):5970-5.
105. Lönnig, Wolf-Ekkehard (2004). «Dynamic genomes, morphological stasis, and the origin of irreducible complexity». Max-Planck-Institut for Plant Breeding Research. Consultado em 9 de setembro de 2017
106. Lopes RJ (2015). Afobação faz cientistas classificarem fósseis de símios como hominídeos. Folha de S. Paulo. Disponível em: http://m.folha.uol.com.br/ciencia/2015/09/1676679-no-afa-cientistas-tem-classificado-fosseis-de-simios-como-hominideos.shtml?mobile
107. Lynch M (2010). Rate, molecular spectrum, and consequences of human mutation.Proc Natl AcadSci U S A. 107(3):961-8.
108. Marchant CJ (1963). Corrected chromosome numbers for Spartina x townsendii and its parent species. Nature. 199:929.
109. Marsh FL (1941). Fundamental Biology. Lincoln, NE: Published by the Author.
110. Marsh FL (1978). Variation And Fixity Among Living Things. A New Biological Principle.CRS Quarterly. 15(2):115-118.
111. Martens K (1997). Speciation in ancient lakes. Trends EcolEvol. 12(5):177-82.
112. Mendonça de Souza, S. M. F., Rodrigues-Carvalho, C., Silva, H. P., & Locks, M. (2006). Revisitando a discussão sobre o Quaternário de Lagoa Santa e o povoamento das Américas: 160 anos de debates científicos. In H. P. Silva & C. Rodrigues-Carvalho (Eds.), Nossa origem. O povoamento das Américas – visões multidisciplinares (pp. 19–43). Rio de Janeiro: Viera & Lentz.
113. Minoletti, Fabrice; Hermoso, Michaël; Gressier, Vincent (2009). «Separation of sedimentary micron-sized particles for palaeoceanography and calcareous nannoplankton biogeochemistry». Nature Protocols. 4 (1): 14–24. ISSN 1750-2799. PMID 19131952. doi:10.1038/nprot.2008.200
114. Mora C, Tittensor DP, Adl S, Simpson AG, Worm B (2011). How many species are there on Earth and in the ocean? PLoS Biol. 9(8):e1001127.
115. Morrell V (1997). Catching Lizards in the Act of Adapting. Science. 276(5313):682-683.
116. Nachman MW, Crowell SL (2000). Estimate of the Mutation Rate per Nucleotide in Humans. Genetics. 156(1):297-304.
117. Nagalingum NS, et al (2011). Recent Synchronous Radiation of a Living Fossil. Science. 334 (6057): 796-799.
118. NASA Astrobiology Institute». nai.nasa.gov. Consultado em 24 de setembro de 201
119. Prüfer K, et al (2013). The complete genome sequence of a Neanderthal from the Altai Mountains. Nature. 505:43-49.
120. ReMine WJ (1993). The Biotic Message. St Paul, MN: St Paul Science.
121. Reinhard Junker, Siegfried Scherer: Evolution – Ein kritisches Lehrbuch. Gießen, ISBN 3-921046-10-6
122. Ridley M (2006). Evolução. 3. ed. Porto Alegre: Artmed. 752p.
123. Robinson DA, Cavanaugh DP (1998). A quantitative approach to baraminology with examples from the primates. CRSQ Quarterly. 34:196–208.
124. Robinson DA (1997). A mitochondrial DNA analysis of the testudineapobaramin. Creation Research Society Quarterly. 33:262–72.
125. Romer AS (1966). Vertebrate Paleontology. 3rd Ed. Chicago: University of Chicago Press, pp. 347–396. In: Denton M. Evolution: A Theory in Crisis. Bethesda: Adler & Adler, 1986, p. 202.
126. Sadava D, Heller HC, Orians GH, Purves WK, Hillis DM (2009). Vida: A Ciência da Biologia. 8. Ed., v. 2. Porto Alegre: EditoraArtmed.
127. Sadler, Peter M. (1 de setembro de 1981). «Sediment Accumulation Rates and the Completeness of Stratigraphic Sections». The Journal of Geology. 89 (5): 569–584. ISSN 0022-1376. doi:10.1086/628623
128. Sanford, John & Baumgardner, John & Brewer, Wesley & Gibson, Paul & Remine, Walter. (2008). Using Numerical Simulation to Test the Validity of Neo-Darwinian Theory.
129. Sanford John C. (2014). Genetic Entropy. 4. ed. New York: FMS Publications.
130. Scherer S (1993). Typen des Lebens. Berlin: Pascal.
131. Schwartz JH, Tattersall I (2015). Defining the genus Homo. Science. 349(6251):931-2.
132. Senter P (2010). Using creation science to demonstrate evolution: application of a creationist method for visualizing gaps in the fossil record to a phylogenetic study of coelurosaurian dinosaurs. Journal of Evolutionary Biology. 23:1732–1743.
133. Sepkoski, J. J. (1993). «Ten years in the library: new data confirm paleontological patterns». Paleobiology. 19 (1): 43–51. ISSN 0094-8373. PMID 11538041
134. Sepkoski, David (2013). «Towards "a natural history of data": evolving practices and epistemologies of data in paleontology, 1800-2000». Journal of the History of Biology. 46 (3): 401–444. ISSN 0022-5010. PMID 22956055. doi:10.1007/s10739-012-9336-6
135. Shabalina SA, Ogurtsov AY, Kondrashov VA, Kondrashov AS (2001). Selective constraint in intergenic regions of human and mouse genomes. Trends Genet. 17(7):373-6.
136. Schopf JW. 1992. Patterns of Proterozoic microfossil diversity: an initial, tentative, analysis. In: Schopf and Klein, p 529-552; (b) Walter MR, Grotzinger JP, Schopf JW. 1992. Proterozoic stromatolites. In: Schopf and Klein, p 253-260
137. Shikano S, Luckinbill LS, Kurihara Y (1990). Changes of traits in a bacterial population associated with protozoal predation. Microbial Ecology. 20:75-84.
138. Snelling A (1997). Sedimentation experiments: Nature finally catches up! Journal of Creation. 11(2):125-126.
139. Souza Jr NN (2008). Uma breve história da terra. Brasília, DF: Sociedade Criacionista Brasileira.
140. Spray, John G.; Kelley, Simon P.; Rowley, David B. (12 de março de 1998). «Evidence for a late Triassic multiple impact event on Earth». Nature (em inglês). 392 (6672): 171–173. ISSN 0028-0836. doi:10.1038/32397
141. Teresi D (2011). Discover Interview: Lynn Margulis Says She's Not Controversial, She's Right. Discover Magazine, p. 68.
142. Trivedi BP (2000). Island mice may evolve faster: From one species to six. Genome News Network. Disponível em: http://www.genomenewsnetwork.org/articles/04_00/island_mice.shtml
143. Van Bocxlaer B, Hunt G (2013). Morphological stasis in an ongoing gastropod radiation from Lake Malawi. PNAS. 110(34):13892-13897.
144. Voje KL (2016). Tempo does not correlate with mode in the fossil record. Evolution. 70(12):2678-2689.
145. Xie, Xufei; Chen, Zhongjing; Peng, Xingyu; Yuan, Xi; Zhang, Xing; Gorini, Giuseppe; Cui, Zhiqiang; Du, Tengfei; Hu, Zhimeng (October 2014). «Neutron emission measurement at the HL-2A tokamak device with a liquid scintillation detector». The Review of Scientific Instruments. 85 (10). 103506 páginas. ISSN 1089-7623. PMID 25362392. doi:10.1063/1.4896120
146. Weyrich A, et al (2016). Paternal intergenerational epigenetic response to heat exposure in male Wild guinea pigs. Mol Ecol. 25:1729-1740.
147. Whitmore J (2013a). Temporal Patterns in 'Living Fossils'. In: 4th Annual Research and Scholarship Symposium. Cedarville, OH. Anais... Cedarville, OH: Cedarville University. Disponível em: http://digitalcommons.cedarville.edu/research_scholarship_symposium/2013/poster_presentations/22/
148. Whitmore J (2013b). What about living fossils? Capítulo 12, pp.143-150. In: Ham K (Ed.). The New Answers Book, v. 4. Green Forest: Master Books.
149. Wills, Matthew A.; Briggs, Derek E. G.; Fortey, Richard A. (1994/04). «Disparity as an evolutionary index: a comparison of Cambrian and Recent arthropods». Paleobiology. 20 (2): 93–130. ISSN 0094-8373. doi:10.1017/S009483730001263X
150. Williamson PG (1981). Morphological Stasis and Developmental Constraint: Real Problems for Neo-Darwinism. Nature. 294:214-2015.
151. Wilmer JW, et al (2011). Catastrophic Floods May Pave the Way for Increased Genetic Diversity in Endemic Artesian Spring Snail Populations. PLoS One. 6(12):e28645.
152. Wilson G (2010). Classic Multidimensional Scaling isn’t the Sine Qua Non of Baraminology. Answers in Genesis. Disponível em: https://answersingenesis.org/creation-science/baraminology/classic-multidimensional-scaling-and-baraminology/
153. Wise KP (1989) The Estimation of True Taxonomic Durations from Fossil Occurrence Data; University Microfilms, 1989.
154. Wise KP (1990). Baraminology: a young-earth creation biosystematic method. In: Walsh RE (Ed.). Proceedings of the Second International Conference on Creationism. Volume II, Technical Symposium, pp. 345–360. Creation Science Fellowship.
155. Wise KP (1992). Practical baraminology. Creation ex Nihilo Technical Journal. 6(2): 122–37.
156. Wise KP (2010) Relatório Gmail [8.7 milion x 300.000]. Mensagem recebida por <sodre.siman@gmail.com> em 13 out. 2015
157. Wittke, James H.; Weaver, James C.; Bunch, Ted E.; Kennett, James P.; Kennett, Douglas J.; Moore, Andrew M. T.; Hillman, Gordon C.; Tankersley, Kenneth B.; Goodyear, Albert C. (4 de junho de 2013). «Evidence for deposition of 10 million tonnes of impact spherules across four continents 12,800 y ago». Proceedings of the National Academy of Sciences of the United States of America. 110 (23): E2088–2097. ISSN 1091-6490. PMID 23690611. doi:10.1073/pnas.1301760110
158. Wood TC, Cavanaugh DP (2001). A baraminological analysis of subtribe Flaveriinae (Asteraceae: Helenieae) and the origin of biological complexity. Origins. 52:7-27.
159. Wood TC, Wise KP, Sanders R, Doran N (2003). A Refined Baramin Concept. Occas. Papers of the BSG. 3:1-14.
160. Wood TC (2001). BDIST software, v. 1.0. Center for Origins Research and Education, Bryan College. Distributed by the author.
161. Wood TC (2002). A baraminology tutorial with examples from the grasses (Poaceae). Journal of Creation. 16(1):15–25.
162. Wood TC (2005). Visualizing baraminic distances using classical multidimensional scaling. Origins. 57:9-29.
163. Wood TC (2010). Baraminological Analysis Places Homo habilis, Homo rudolfensis, and Australopithecus sediba in the Human Holobaramin. Answers Research Journal 3:71–90.
164. Wood TC (2011). Using creation science to demonstrate evolution? Senter’s strategy revisited. Journal of Evolutionary Biology. 24(4):914–918.
165. Woodmorappe J (2000). The Karoo vertebrate nonproblem: 800 billion fossils or not. CEN Technical Journal. 14(2):47-49.
166. Woodruff DS (1980). Evolution: The Paleobiological View. Science. 208(4445):716-717.
167. Zhang, Y. P.; Yang, J. W.; Liu, Yi; Fan, T. S.; Luo, X. B.; Yuan, G. L.; Zhang, P. F.; Xie, X. F.; Song, X. Y. (June 2016). «Development of the radial neutron camera system for the HL-2A tokamak». The Review of Scientific Instruments. 87 (6). 063503 páginas. ISSN 1089-7623. PMID 27370450. doi:10.1063/1.4953109
168. Zellner, Nicolle E. B. (1 de setembro de 2017). «Cataclysm No More: New Views on the Timing and Delivery of Lunar Impactors». Origins of Life and Evolution of Biospheres (em inglês). 47 (3): 261–280. ISSN 0169-6149. doi:10.1007/s11084-017-9536-3
169. Zimmerman EC (1960). Possible evidence of rapid evolution in Hawaiian moths. Evolution. 14:137-138.
Martini, I. Peter; Baker, Victor R.; Garzón, Guillermina (5 de março de 2009). Flood and Megaflood Processes and Deposits: Recent and Ancient Examples (Special Publication 32 of the IAS) (em inglês). [S.l.]: John Wiley & Sons. ISBN 9781444304305 Disponível em https://books.google.com.br/books?hl=pt-BR&lr=&id=57wX5svvh8AC&oi=fnd&pg=PA3&ots=yPHV0d7eME&sig=cF1sJVvf1prTkt05DpnKRT2tqIw#v=onepage&q&f=false acessado em 16/12/2017
Sodré, GBN.; Souza Junior.; 2010 «Tempo de Formação: do Cambriano ao Pleistoceno | pt-brAdam's Apple». www.adamsapple.com.br. Consultado em 29 de novembro de 2017
Saitta, Evan T.; Rogers, Christopher S.; Brooker, Richard A.; Vinther, Jakob (1 de outubro de 2017). «Experimental taphonomy of keratin: a structural analysis of early taphonomic changes». PALAIOS: 647–657. ISSN 0883-1351. doi:10.2110/palo.2017.051
Axe, Douglas D.; Dixon, Brendan W.; Lu, Philip (4 de junho de 2008). «Stylus: a system for evolutionary experimentation based on a protein/proteome model with non-arbitrary functional constraints». PloS One. 3 (6): e2246. ISSN 1932-6203. PMID 18523658. doi:10.1371/journal.pone.0002246«Signature in the Cell « ASA Book Discussion». www.asa3online.org (em inglês). Consultado em 13 de outubro de 2017
Weber BH (1998) Emergence of life and biological selection from the perspective of complex systems dynamics. In: Van De Vijver G, Sathe SN, Delpos M (eds) Evolutionary Systems: Biological and Epistemological Perspectives on Selection and Self-Organization, pp 59–66. Kluwer, Dordrecht.
Kauffman SA (1993) The Origins of Order: Self-Organization and Selection in Evolution. Oxford University Press, New York.
Morowitz H (1992) Beginnings of Cellular Life: Metabolism Recapitulates Biogenesis. Yale University Press, New Haven.
Weber BH (2010) What is Life? Defining life in the context of emergent complexity. Origins of Life and Evolution of Biospheres 40: 221–229.
Schmitz, B. , Tassinari, M. & Peucker-Ehrenbrink, B. Uma chuva de condritas comuns no Ordoviciano adiantado . Planeta Terra. Sci. Lett. 194 , 1 - 15 ( 2001 ).
Schmitz, B., Tassinari, M. & Peucker-Ehrenbrink, B. A rain of ordinary chondrites in the early Ordovician. Earth Planet. Sci. Lett. 194, 1–15 (2001).
NATURE ASTRONOMY 1, 0035 (2017) | DOI: 10.1038/s41550-016-0035 | www.nature.com/natureastronomy 1
LETTERS
PUBLISHED: 23 JANUARY 2017 | VOLUME: 1 | ARTICLE NUMBER: 0035
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
Rare meteorites common in the Ordovician period
Rare meteorites common in the Ordovician period (PDF Download Available). Available from: https://www.researchgate.net/publication/312647580_Rare_meteorites_common_in_the_Ordovician_period [accessed Nov 21 2017]. «Rare meteorites common in the Ordovician period (PDF Download Available)». ResearchGate (em inglês). Consultado em 21 de novembro de 2017
HISTORIA DA TNI COMEÇA em 2017 COM A PUBLICAÇÃO DE UM ARTIGO SOBRE PMS - (paradoxo da repetição das formas fósseis)
"Assustado com a reação busquei mais fontes que corroborassem com meu artigo para defendê-lo desta tempestade de críticas e um dos primeiros artigos que encontrei foi do grande cientista alemão Wolf-Ekkehard Lönnig e fiquei digamos aliviado ao ler quase as mesmas observações em "Dynamic Genomes" . Raciocinei que se tão grande cientista (índice h-16 scopus) pensou as mesmas coisas, eu deveria estar num caminho já trilhado e não em uma novidade insipiente que a academia brasileira alardeou". ***(Assista ao VIDEO com uma introdução geral ao artigo).
A Polêmica publicação comentada em outros sites :
Como começou a TNI
Tudo começou com a publicação de um artigo falando sobre PMS- Paradoxo da estase morfológica sendo interpretada como clara evidencia de sepultamento de todas as espécies ancestrais da terra num só momento, pois se a evolução é fato, e é , ela nunca deixaria espécies em estase morfológica por milhões de anos .
Esta publicação finalizava levantando a necessidade de estudos sobre hipótese de chuva de asteroides com seus efeitos sedimentares capazes de enterrar toda fauna terrestre e assim preservar em estase e repetições tais populações sepultadas.
Mais tarde ao defrontar com anacronismos percebemos possiveis efeitos de "envelhecimento" de rochas ao acelerarem decaimento radioativo por algum mecanismo elétrico, plasma, altas temperaturas envolvidas num bombardeamento de asteroides, quem sabe poder desencadear até mesmo possíveis colisões de íons pesados, justificando e explicando uma série de dados anacronicos, radioativos, anomalias , temporalidades, geoquimica e de altíssima temperatura , que imperam as vezes como "mistérios cientificos" na atualidade.
No primeiro artigo, (que foi criticado era quase apenas em relação a categoria da revista em que foi publicado e não ao seu conteúdo), porém o conteúdo revelou-se ser relevante até pelo fato de chamar tanto atenção, mesmo sendo publicado numa revista de pouco prestigio.
No artigo listamos alguns padrões fósseis como:
1. Padrão fóssil de ter repetições de formas e repetição taxonômica fóssil em estratos geológicos diferentes
2. Padrão fossil de pobreza taxonômica comparada a grande diversidade taxonômica biológica hoje
3. Praticamente termos o mesmo número de padrões corpóreos fósseis com os padrões corpóreos existentes vivos (ou seja, aumenta a biodiversidade mas não aumenta numero de padrões corpóreos) - referenciado no trabalho com troca de e-mail entre eu e e um doutor em paleontologia tendo tese de "estatistica fossil" defendida em Harvard, sob orientação de Stephen Jay Gould.
Estes padrões estão postos como evidencia de sepultamento de todos os seres vivos da terra (justificando repetições) representados pelas espécies mães ancestrais "baraminológicas" ou como Darwin citava de outros autores "espécies originais" (justificando a pobreza taxonômica) como tipos básicos ancestrais morfologicamente estáveis numericamente (justificando os limites evolutivos em torno dos padrões morfológicos das mesmas)
PMS - Uma briga antiga na academia
"Em 1944 (Simpson, 1944) fez uma meta-análise e concluiu a partir de 58 estudos publicados sobre padrões de especiação no registro fóssil que 71% apresentavam estase (e permanência) morfológica. A este respeito Michael Benton declarou: "parece claro, então, que a estase é comum, e isso não foi previsto a partir de estudos genéticos modernos" (Benton, 2009; Michael Oards, 2017), que preveem imensa variação de formas a partir do comportamento plástico dos seres vivos observados sobretudo desde Charles Darwin.
Por quase uma década mesmo sem ter o conhecimento de muitas destas pesquisas sobre a anomalia da "estase morfológica" , defendia o que chamava de "Paleontologia em T" , destacando o contraste entre os achados fósseis se repetindo em permanência de formas, com a altíssima variabilidade de formas recentes na "atualidade", e me baseei nesta época mais na diferença taxonômica e depois também no contraste entre fósseis repetidos em suas mesmas formas com seres vivos plásticos em seu comportamento usual (Lauvoué, 2009)
Anos mais tarde um cientista experiente em publicações (Everton Fernandes) me contactou para que publicássemos juntos algum dos muitos assuntos que me envolvi. Sugeri este tema (assunto que me motiva) com o título: "A Especiação em Tempo Real, em Tempo Histórico-Arqueológico e sua Ausência em Tempo Geocronológico" sendo publicado em inglês "Speciation in real time and historical-archaeological and its absence in geological time" publicado pela Academia Publishing sob DOI: 10.15413/ajsr.2017.0413 em maio de 2017. PDF com revisão em inglês
Durante a construção do artigo houveram varias descobertas e uma delas o Everton descobriu que, o que eu defendia fazia vários anos, como "Paleontologia em T" enfatizando a explosão taxonômica de muitas espécies no presente, era chamado na literatura de "estase morfológica" que destaca mais um padrão fóssil não só da pobreza taxonômica, mas da pobreza morfológica também, destacando a imobilidade de variação de formas e sua repetição morfológica e de espécies nos estratos geológicos.
Os artigos que tratam da "estase morfológica" são caracterizados por justificativas do tipo "ad hoc" .
"ad hoc significa a adição de hipótese(s) estranha(s) a uma teoria para salvá-la de ser falseada. Hipóteses ad hoc compensam anomalias não previstas pelas teorias em sua forma original, ainda não modificada. Cientistas são frequentemente céticos sobre teorias que as apresentam frequentemente, não suportando ajustes para sustentá-las. Hipóteses Ad hoc são frequentemente características de pseudocientistas. Muito da produção científica reside sobre a modificação de hipóteses existentes ou teorias mas estas modificações são distintas de hipóteses ad hoc no que as anomalias são explicadas propondo um novo significado daquilo que é real e observável".
Gardner, Martin. The Whys of a Philosophical Scrivener [Os Porquês de Um Escrevente Filosófico] (New York: Quill, 1983).
Gould, Stephen Jay. Ever Since Darwin [Desde Darwin] (New York: W.W. Norton & Company, 1979).
Em vez de simplesmente admitir o problema e buscar outra teoria histórica dos seres vivos, uma vez que torna-se impossível enxergar o contraste de formas permanentes no registro fóssil com o comportamento não-permanente em mesmas formas na biodiversidade atual (super diversificada e se variando morfologicamente), até mesmo em mesmas especies quando sob quaisquer mudanças ambientais, pois "nenhum organismo ignora seu ambiente"(Ezard et e al, 2016), as publicações procuram uma forma de salvar a teoria e manter o mesmo histórico por mais que os dados falem contra.
Mas enfim de posse das palavras chaves "estase morfológica" foi bem mais fácil encontrar inúmeros outros artigos científicos tratando da questão até encontrar a expressão (contendo em si mesma) a flagrante e escandalosa admissão do "maior problema da evolução" (Mayer): PMS (Paradox of Morphological Stasis) "o paradoxo da estase morfológica" e são estas as palavras chaves do nosso antigo artigo.
A idéia do nosso artigo anterior era apenas demonstrar a anomalia das mudanças de formas (esperadas mas não achadas) e do padrão da repetição de formas e repetição taxonômica fóssil (mesmas espécies se repetindo até em estratos diferentes) como evidencia de sepultamento de seres ancestrais iniciais básicos de todo planeta, enquanto Wolf-Ekkehard Lönnig destaca os mesmos como surgidos prontos na coluna geológica como uma necessidade a sobrevivência uma vez que é necessário uma "complexidade irredutível" (Behe) para tal.
Logicamente que isso tinha sérias implicâncias usando a própria "evolução fato" para desconstruir o suposto histórico evolucionista das especies por meio da apresentação do PMS , mas também apoiava a idéia criacionista com 200 anos de altíssima rivalidade , de uma catástrofe mundial capaz de sepultar milhares de populações surgidas prontas num ambiente comum e por isso estarem repetidas em suas mesmas formas nos achados fosseis, já que até mesmas espécies tendem a mudar suas formas (quase todas) nas alterações de ambiente.

O sepultamento de todas as populações da terra é uma interpretação falseável para o PMS ( padrão de repetição de mesmas formas fosseis).
O que causou os dilúvios e catástrofes ígneas da terra tem relação com quatrilhões de pedras ainda pontiagudas e sem desgaste mesmo aquelas sob forte impacto de águas nas cachoeiras
Quando a academia brasileira soube da publicação houve drástica reação! Ameaçaram a terceira autora de perder a bolsa de pós doutorado em evolução (era a segunda perseguição frontal que a garota recebia) e mesmo sem respeitar o estado "grávido" dela, muito provavelmente a pressionaram para que solicitasse a remoção de seu nome, o que a revista se recusou fazê-lo. Na época tive então que declarar a isenção ao máximo de sua participação (até porque comigo não sei quantificar , mas acredito que foram poucas mesmo, não sei com o segundo autor) depois fizeram uma tempestade de protestos contra o artigo nas redes sociais, onde tentei quase em vão , discutir o assunto do artigo e chamar para o debate quem se dispusesse.
Podemos observar como destaco no artigo abaixo, que existem confissões flagrantes do tipo "ato falho" (Freud) como julgo ser a análise do conservadorismo morfológico contrastado a plasticidade genética do "Pantodon buchholzi" feita por Lavoué: "estudar o conservadorismo morfológico a longo prazo é difícil nos sistemas contemporâneos, porque poucas linhagens animais existentes são conhecidas por permanecerem morfologicamente estáticas em relação às escalas de tempo geológicas". (Lavoué et al, 2011).microceb@hotmail.com
Apesar de nosso artigo destacar pontos falseáveis, podemos supor que isto se deu porque a academia mundial principalmente das áreas de geologia e biologia, é bem conhecida pelo seu comportamento repleto de repulsa para com artigos científicos criacionistas e do design inteligente (conforme um dos mais assistidos documentários do mundo "Expelled: No Intelligence Allowed"), o qual descreve dezenas de casos onde cientistas respeitados foram boicotados, mandados embora, prejudicados por apoiarem de alguma forma o criacionismo ou o design inteligente, e previsivelmente reagiu repudiando o artigo devido o caráter nitidamente a favor do "criacionismo".
Era até esperada esta reação por conta da nossa "afronta e audácia" de publicar dentro do terreno protegido e vigiado pelo que o Dr. Eberlin (um dos maiores bioquímicos da atualidade) chama de "patrulha darwinista", que deixou escapar a publicação em jornal internacional de idéias a favor do seu arqui-rival histórico e político (criacionismo) que desde o iluminismo tratou de considerar indistintamente separação política entre igreja e Estado (laicismo político) como válidos também na educação (laicismo educacional), tratando de simplesmente como Freud sinalizou ao propor substituir as versões da religião para reinterpretar a realidade de uma nova maneira por considerar as descrições dela doentias e retrógradas.
Proibir assuntos como criador, planejador, design inteligente, em sala de aula e varrer para debaixo do tapete da observação a existência de um codificar de códigos, um informante para informação, um ajustador de "Fine-tuning", um legislador de leis, e declarar que o que o pais da ciência chamavam de principio básico (Criador) como algo algo inexistente, se tornou a tarefa "missionaria" libertadora de muitos acadêmicos, criando assim, um exército militante e formador de opinião de ateus e céticos junto a uma distância gigantesca entre o pensamento dos pais da ciência moderna e a própria ciência moderna, que agora, sob os moldes da camisa de força "naturalista" sufocando assuntos que evoquem temas arqueológicos e religiosos, substituindo-os por mera ficção científica (sopa primordial, panspermia, biopoeise, etc) se tornando ciência moderna preconceituosa quase apenas em relação a temas relacionados ao criacionismo antigo.
Bem ao contrário disso os pais da ciência como Isaac Newton foram estimulados cientificamente pelas ideias religiosas de haver um sistematizador e legislador para encontrar leis, como vemos em seus escritos:
Em menos de uma semana . assim que se descobriu meu artigo no "researchgate" Houveram provavelmente dezenas de milhares de reações acadêmicas no Brasil e algumas internacionais como de uma autoridade britânica de um comitê para o "progresso" da ciência.
Na grande maioria das reações não houve apontamento ou demonstração de entendimento do assunto , houveram 3 ou 4 cientistas que se deram ao trabalho de debater numa página destinada a isso , mas a maioria teve como saída apenas atacá-lo por ser criacionista e por ter sido publicado numa revista da lista das possíveis "predatórias" apesar que o corpo editorial da mesma possuir boas notas B3, B4 conforme lista de 2017 pela agência oficial CAPES que apoia pesquisadores brasileiros.
Assustado com a reação busquei mais fontes que corroborassem com meu artigo para defendê-lo desta tempestade de críticas e um dos primeiros artigos que encontrei foi do grande cientista alemão (não poderia ser um brasileiro? ) Wolf-Ekkehard Lönnig e fiquei digamos aliviado ao ler quase as mesmas observações em "Dynamic Genomes" . Raciocinei que se tão grande cientista pensou as mesmas coisas eu não estaria tão absurdamente errado como a academia brasileira alardeava.
Encorajado a pesquisar mais busquei antecipar objeções, e dar resposta mais completa, abrangente e eficiente para satisfazer questões levantadas pelo mesmo artigo. Comecei fazendo acréscimos e revisões para antecipar as críticas, mas pouco tempo a revisão se transformou não somente num novo artigo, mas em uma teoria, com centenas de elementos novos ou mais desenvolvidos, centenas de artigos citados a mais, onde o ponto principal que era " o paradoxo da estase morfológica"observada como anomalia sem qualquer harmonia com a teoria de Darwin (observado por Darwin e antes dele), se tornou ponto lateral e coadjuvante junto com dezenas de outras hipóteses algumas testadas e outras sugeridas em estudo, formando um novo modelo geocronológico de impactos que devido algumas destas hipóteses já estarem testadas e devidamente publicadas denominamos "Teoria Neocatastrofista de Impactos" com a qual percebemos que os diversos problemas atuais da historia da terra inclusive a PMS (Paradox of Morphological Stasis), funcionam neste novo modelo/teoria como peças de uma quebra-cabeças permitindo uma visão cada vez mais harmônica com a realidade.
Recentemente encontrei experiente cientista seguindo mesma linha (Michael Oards), além dos muitos citados neste trabalho como o Dr. Walter Brown, Loening do Max Plank, Dr. Nahor Souza Junior que foi quem me deu os fundamentos da geologia catastrofista e o primeiro a falar para mim que os impactos provavelmente eram "o segredo" e foi a partir de um mega debate de 9000 réplicas e tréplicas, com 4 professores de geologia durante 1 ano e meio sob assistência do mesmo, que percebi um pouco que os impactos eram o ponto chave para a compreensão da historia da terra e sobretudo do fanerozoico, os quais complementam dentro de suas especialidades e pontos observados, a confirmação de que estamos apenas um pouco mais acima dos efeitos dos mesmos.
Por fim busquei e ainda busco uma centenas de revisores, críticos, colaboradores e co-autores entre os quais cito Hector Lutero, que foi quem percebeu geniosamente a hipótese de tendência de relação de diâmetro do bólido com idade (Figura 1), Dan de Barna, que me forneceu um capítulo inteiro de seu livro citandio o famoso cientista "Alvares" , Roberto Carlos Machado que forneceu relatos arqueológicos de catástrofes e tem pesquisado sobre cinturão de asteroides, e fui agregando alguns pontos importantes e revisando outros, podendo oferecer agora com uma certa expectativa positiva, uma previsão de trabalho ao público em geral, que mesmo possuindo diversos pontos que ainda necessitam de maiores estudos e demonstrações, ofereço para que a discussão do mesmo e sua exposição, possa melhorar ou criar novos projetos de pesquisa atrelados, que dignifiquem cada vez mais a ciência dos fatos, nossas descobertas , estes novos autores e suas novas ou antigas publicações, com novas observações e olhares ; e porque não mencionar também, para que dignifiquem sobretudo e sobre todos ao nosso Criador.
A todos uma boa leitura!
Sodré Neto
geologiaatual@gmail.com
GLOSSARIO
Paradoxo da Estase Morfológica - Permanência de mesma ou semelhante forma durante os achados do registro fóssil nos estratos e camadas geológicas. Esta permanência é precedida por surgimento pronto (sem etapas evolucionarias achadas antes nem lateralmente no sentido de ligar evolucionariamente um padrão morfológico fóssil ao outro (disparidade ou diferenciação de padrões morfológicos aumentada no registro fóssil) , bem como extinção da forma no registro fóssil , e as vezes reaparecimento daquela forma extinta , da espécie ou forma padrão, viva hoje .
Geocronologia - A geocronologia convencional seria a datação "absoluta" adotada para datar as camadas geológicas como se fossem períodos de tempo geológicos . Já a geocronologia catastrofista publicou exemplos como o cambriano em artigos de jornais especializados de geologia , se tratar de mega-estratificações geradas por altíssimas taxas sedimentares durante evento catastrófico e rápido de "magnitude global" (Souza, N J, 2001). Esta segunda opção de interpretação dos fatos geológicos explica dobras
|
|
| Back | World Map | North America | South America | Europe | Asia & Russia | Africa | Australia | Earth Impact Database Sorted by : | Name | Diameter | Age | Impact Structures Sorted by Diameter
****This revised diameter is the best estimate for the collapsed transient crater diameter (rim-to-rim dimension). Our previous diameters cited maximum damage diameter estimates. There is considerable confusion in the literature regarding the definition of "diameter". In the Earth Impact Database, we are striving to cite the collapsed transient crater value where possible. This can affect the order of size: for example, Sudbury's maximum damage diameter is ~260 km (as defined by the outermost ring diameter), while that of Chicxulub is ~240 km. However, the rim-to-rim diameter of Sudbury is less than that of Chicxulub's (130 versus 150 km, respectively). |
